
Evolutionary theory asserts that today’s plants and animals developed from far different ancestral life-forms. The characteristics of populations have changed with time, to produce new species. These changes are believed to be relatively gradual. Scientists believe that a new species may take thousands of generations to become clearly differentiated from its original form. Major transitions, such as going from fish to amphibians, or reptiles to mammals, must have involved millions of years and a whole range of intermediate forms. We intuitively expect to see evidence of these intermediate forms in the fossil record.
Mainstream science says that plenty of such transitional fossils have been found. Young Earth (YE) creationists and Intelligent Design advocates disagree. They cite what Darwin himself wrote in the Origin of Species (6th Ed., pp. 341-342), “The number of intermediate varieties, which have formerly existed must be truly enormous. Why then is not every geological formation and every stratum full of such intermediate links? Geology assuredly does not reveal any such finely graduated organic chain; and this, perhaps is the most obvious and serious objection which can be urged against the theory.”
Thus, transitional fossils are a significant part of the evidence for or against evolution. It is necessary to have a correct understanding of the nature of transitional fossils, in order to properly evaluate the physical evidence. Four factors which impact the nature of fossil intermediates are described below.
First, a little vocabulary: living organisms are grouped together in various levels. From highest level (most inclusive) to lowest, these are: domain, kingdom, phylum, class, order, family, genus, and species. Below species there are strains or sub-species. All animals are grouped in one kingdom, and all plants in another. These are mainly human constructs forced onto the messy biological world, and scientists can legitimately differ on how any particular organism should be classified. A common definition for a plant or animal “species” is the largest group of organisms capable of interbreeding and producing fertile offspring.
See Whale Origins: A Test Case for Evolution for discussion of fossil and genetic evidence regarding the evolution of whales.
Four Key Factors Governing the Transitional Fossil Record
(1) The fossil record is inherently very sparse. Very, very few of all the organisms that have died in past eons become fossilized. As we can observe today, nearly all carcasses rot or are eaten by scavengers rather than being buried intact in rock layers. Of the remains that do get fossilized, many are later eroded away if the rocks in which they are embedded are raised above sea level. If these rocks become deeply buried, the fossils can become smeared beyond recognition in metamorphic transformations. This is even more of an issue for older rock layers, such as the Paleozoic era (Cambrian through Permian periods), since they have had more time to be either raised up to the surface for erosion or to be buried more deeply. Also, of all the potential fossil-bearing rocks, only a small fraction is available in surface exposures for paleontologists to examine.
As Wikipedia points out, “The number of species known through the fossil record is less than 5% of the number of known living species, suggesting that the number of species known through fossils must be far less than 1% of all the species that have ever lived.” No fossils have yet been found for about a third of the 30+ phyla of living animals. Occasionally (e.g. once every 15 million years or so) we find a rock formation such as the Burgess Shale where conditions were just right to preserve a rich assemblage of fossils (including many soft-bodied animals) in that locale at that snapshot in time. These “Lagerstatte” are the exceptions which prove the rule: they confirm that the ancient seas were teeming with diverse life-forms, but in most times and in most places (i.e. apart from these very rare fossil-rich formations), these organisms simply did not become preserved as recognizable fossils.
The Coelacanth order of fishes furnishes a classic example of the fickleness of the fossil record. These fish were once widespread in the ancient seas. Coelacanths peaked in the fossil record about 240 million years ago, and then declined. The most recent known fossil dates back to about 80 million years ago. It was thought that they had become extinct. In 1938, however, a live coelacanth was discovered in the Indian Ocean. Since then a number of others have been caught. Unless we are prepared to claim that an Intelligent Agent supernaturally re-created these modern coelacanths, we must acknowledge that some population of these fish has existed for the past 80 million years but without leaving a trace in the fossil record.
We should expect to observe many gaps like this in the fossil record. Here is a list of other “Lazarus taxa” which disappear from the fossil record for millions of years, but appear again later.
(2) New species tend to develop in small, isolated populations. The arithmetic of basic population genetics shows that it is more difficult for new genetic mutations to become established in very large populations, than in small populations. This is readily confirmed by laboratory studies. For instance, Perfeito et al. found that new beneficial mutations were much more readily established in small populations of bacteria than in large populations.
Thus, it is far more likely that a new species would develop within a small, isolated population, especially if that population is under some environmental stress that would favor genetic changes. The odds of us finding fossils from that small, localized population is are nearly zero. If the new species becomes more fit than the old species, the new species will expand, and only then is likely to appear in the fossil record. But once a species is widespread and successful in its ecological niche, there will be diminished selection pressure for changes, so fossils of this now well-adapted species are likely to appear for perhaps million of years with showing little change.
(3) A given population can persist for many millions of years with little morphological change. As demonstrated by the longevity of the coelacanth group, a specific type of organism can persist for tens of millions of years with only modest changes. Thus, if we find a fossil of some species in rocks dated as being, say, 100 million years old, it is quite possible that a similar, related species (same genus or family) also existed 110 million years ago, and maybe even 150 million years ago, whether or not we have found these older fossils yet. For instance, the past 80 million years would constitute such a “ghost” lineage for coelacanths.
(4) Evolutionary lineages tend to be “branchy”. Typically what shows up in the fossil record are organisms on the side branches, rather than the directly ancestral ones along the main “trunk” of the family tree. These side branch species often show intermediate features, but are not the actual transitional fossils.
Horse Evolution: Example of Branching Lineage
The evolution of today’s horse (genus “Equus”) from a tiny ancestor is well-supported in the

Source: Professor Donald Levin’s course in BioEvolution at the University of Texas in Austin, copied from http://darwiniana.org/horses.htm.
fossil record. Hyracotherium (sometimes called Eohippus, i.e. “Dawn Horse”) lived in the forests about 55 million years ago. It stood roughly 18 inches (50 cm) tall at the shoulder, resembling a small dog. Its teeth were adapted to eating fruit and soft foliage, and its feet and leg bones were quite different from today’s steeds that thunder across the plains and eat tough grass. The graphic on the right depicts some of the changes in teeth, toes, and overall body size.
The picture below, from the Florida Museum of Natural History, shows how the size and shape of fossil horse skulls has changed with time:

Source: “Fossil Horses” on-line exhibit, by the Florida Museum of Natural History http://www.flmnh.ufl.edu/fhc/Stratmap1.htm
In her article at TalkOrigins on horse evolution, Kathleen Hunt shows a more complete family tree (grouped at the level of genus), with even more branches than depicted in the figures above:

Source: “Horse Evolution” by Kathleen Hunt, at http://www.TalkOrigins.org
Hunt notes a number of clear, graduated changes that appear in this fossil series. For instance, Hyracotherium (“Eohippus”) had only three grinding molars on each side of its jaw, Orohippus had four, Epihippus had five, and Mesohippus had six grinding “cheek teeth.” That looks pretty “transitional” to me. Although a clear line of descent can be drawn between Eohippus and today’s horses, Hunt cautions that the fossils show a complex pattern, not a unidirectional trend:
Horse species were constantly branching off the “evolutionary tree” and evolving along various unrelated routes. There’s no discernable “straight line” of horse evolution. Many horse species were usually present at the same time, with various numbers of toes, adapted to various different diets…Tracing a line of descent from Hyracotherium to Equus reveals several apparent trends: reduction of toe number, increase in size of cheek teeth, lengthening of the face, increase in body size. But these trends are not seen in all of the horse lines. On the whole, horses got larger, but some horses (Archeohippus, Calippus) then got smaller again. Many recent horses evolved complex facial pits, and then some of their descendants lost them again. Most of the recent (5-10 My) horses were three-toed, not one-toed, and we see a “trend” to one toe only because all the three-toed lines have recently become extinct.
In his general article on transitional fossils, Keith Miller offers these observations on the horse lineage:
Interestingly, some critics of evolution view the record of fossil horses from “Eohippus” (Hyracotherium) to Equus as trivial. However, that is only because the intermediate forms are known. Without them, the morphologic distance would appear great. “Eohippus” was a very small (some species only 18 inches long) and generalized herbivore (probably a browser). Besides the well-known difference in toe number (four toes at front, three at back), “Eohippus” had a narrow elongate skull with a relatively small brain and eyes forward in the skull. It possessed small canine teeth, premolars, and low-crowned simple molars. Over geologic time and within several lineages, the skull became much deeper, the eyes moved back, and the brain became larger. The incisors were widened, premolars were altered to molars, and the molars became very high-crowned with a highly complex folding of the enamel.
The significance of the fossil record of horses becomes clearer when it is compared with that of the other members of the order Perissodactyla (“odd-toed ungulates”). The fossil record of the extinct titanotheres is quite good, and the earliest representatives of this group are very similar to “Eohippus“. Likewise, the earliest members of the tapirs and rhinos were very “Eohippus“-like. Thus, the different perissodactyl groups can be traced back to a group of very similar small generalized ungulates.
Expectations for Fossil Lineages
Figure 1 represents a naïve expectation of what the fossil record should look like for the evolutionary family tree encompassing some species A through D. In this figure, there is a single lineage, with the direct ancestors all appearing as fossils. Each earlier form neatly disappears from the fossil record as the next one appears, so there are no overlaps. From the four factors discussed above, it is obvious why Figure 1 is not realistic, yet this is what YE creationists often demand to see.

Figure 2 shows a more realistic fossil lineage. The points of actual divergence (common ancestors) are relatively unlikely to have left fossils. It is more probable that we will find fossils of successful populations on the side branches of the family tree, such as B and C in this figure. This is not due to some weakness in evolution. Rather it is due to the intrinsic nature of speciation and fossil preservation (branching, new species arising in small populations, etc.).

While this diagram shows the branching which is characteristic of evolutionary lineages, it is unrealistic in depicting the rate of morphological change as being roughly constant with time. As noted above, evolutionary changes tend to happen relatively quickly in small populations that do not leave a trace in the fossil record, while large successful populations can continue with only modest changes for many millions of years. (By “relatively quickly” we mean that a new genus might develop in a million years or so for vertebrates, and of course in shorter times for organisms with faster reproductive rates.)
Figure 3 below better represents the type of fossil pattern we expect from what is known about speciation and fossilization:

The thick vertical lines denote the chronological extent of the fossil remains for the four species A–D. There are fossils of B and C which display some characteristics which are intermediate between A and D. However, some of these fossils do not appear in strict morphological order. All of the extant fossils of C predate all the fossils of B, even though the morphological progression suggests that B would be in some sense ancestral to C. Some fossils of D appear at the same time as fossils of C.
YE creationists sometimes claim these “out of order” fossils represent some sort of dire problem for evolution. This claim is false. This type of pattern is exactly what is predicted by the four factors discussed above. Populations like A, B, C, and D that actually appear in the fossil record are likely to be large and stable. But there will always be an earlier time, before these populations expanded (or when geological conditions did not favor preservation), when each of these populations was present but was not represented among the known fossils. It is entirely plausible that a common ancestor between B and C existed sometime before the earliest fossils of C, even though the lineage from that ancestor up to the B fossils has not yet been observed in the rock layers.

A possible family tree for these fossils is shown above in Figure 4. The thick lines denote the observed fossils, with the thin lines denoting inferred lineage relationships.

Another potential family tree for the same fossils is shown above in Figure 5. It shows a slightly different pattern of splits from common ancestors. How can we determine which lineage pattern is a better fit to the fossil evidence?
Arthropods: Example of Fossil Family Tree
There is a whole field of study called cladistics where scientists ponder these questions. They define and analyze various characteristics of physical features in fossils, and also genetic relations among living species. Here is an example of the type of thinking that goes into cladistics, centered on the evolution of arthropods (think: bugs) about 500 million years ago. Dennis Venema at Biologos writes:
All living arthropods have a suite of defining characteristics such as a hard external skeleton (exoskeleton), specialized body segments, and specialized appendages. While these characteristics are useful for defining modern arthropods, these criteria become less useful as we travel back through the evolutionary history of arthropods. The reason is simple – from an evolutionary point of view, one would not expect these different traits to arise as a unit in one fell swoop. Rather, one would expect that these traits would arise over time in the lineage leading to modern day arthropods. [Thus] we would expect to find species in the fossil record that do not have the full suite of “arthropod” characteristics, but only some:

Source: Dennis Venema, “Evolution Basics: The Cambrian Diversification and Assembling Animal Body Plans, Part 1”, at http://www.biologos.org
For example, based on the above phylogeny we might expect to find two groups of “arthropod-like” organisms in the fossil record: species that have only (1) of the three traits (specialized appendages only), as well as a second group (2) with specialized appendages and segments. …these species would represent “transitional forms” in the sense that they have intermediate sets of characteristic features that indicate the steps the arthropod lineage took to achieve the “modern” suite of characteristics.
And in fact, this sort of pattern is reflected in the fossil record. The figure below, from Legg, et al., shows that the ancient fossils are consistent with sequential modifications in appendages and body segments as populations developed from early arthropod-like forms to full modern-type arthropods. Here, modern arthropods (e.g. Hexapoda [insects], and Crustacea [crabs, etc.]) are shown at the bottom, with the earliest evolutionary branching at the top. For instance, Anomalocaris had large compound eyes and hardened specialized appendages, but it did not have a hardened exoskeleton over its whole body. It is considered a transitional “stem” arthropod, not a full-fledged “crown” arthropod. Each point (1, 2, 3, etc.) represents the appearance of a key innovation, such as compound eyes, arthropodized limbs, arthropodization of the trunk exoskeleton, etc.

Source: D.A. Legg et al., Proc. Royal Soc. B, 2012: vol. 279 , pp. 4699-4704
The cladistic relationships in this figure were not just randomly proposed to rationalize the fossils. Nick Matzke described the rigor that went into constructing this diagram:
These researchers, and the previous researchers that they are building upon, identified 580 individual, variable characters, each of which has to be identified, defined, divided up into discrete character states, and encoded. This laborious process had to be repeated for (in this) case 173 fossil [or living] taxa… A lot of fossils are missing a lot of characters – typical and expected in paleontological analyses – but it is still a lot of work. After this, one runs a cladistic or other phylogenetic analysis… and calculates support statistics. …Almost any biological dataset typically has extremely statistically significant tree signal, and this is true whether or not it agrees precisely with other analyses, and whether or not all relationships of interest to the researcher are precisely resolved with high support.
Fish-To-Tetrapods: A Missing Link Is Found?
400 million years ago, the seas were teeming with various types of fish, most of which lived entirely underwater, and had fairly flimsy fins. By about 365 million years ago, fossil evidence exists for primitive tetrapods, such as Acanthostega and Ichthyostega. These creatures had four limbs which could support some weight and assist in moving around in shallow water, although they probably could not readily walk around on dry land. How did fish evolve into tetrapods? Besides the development of limbs, the bones of the head had to change from two parts to one, and the solid head/body connection in a fish was replaced by a flexible neck.
The leading candidates among fishes for tetrapod ancestry are the “lobe-finned” fishes. The bottom fins of these fishes are arranged in pairs and supported by internal bones, which would facilitate the evolution of these fins into legs. Also, the lobe-finned fishes of the Devonian period were already able to breathe air, when necessary, via “spiracles” in their skulls.
Various lobe-finned fish fossils have long been known, where the skeletal structures of the fins include primitive versions of the bones found in the limbs of later, terrestrial animals. Examples include Eusthenopteron and Panderichthys, whose remains have been dated to about 380 million years ago. While these fossils hint at evolutionary pathways toward tetrapods, these are still very much like regular fishes. It would be gratifying to discover a fossil species with more pronounced tetrapod-like features.
In 1999 a team of scientists led by Neil Shubin of the University of Chicago set out to find such a transitional fossil. Since the main fish-tetrapod fossil gap was in the 363-380 million year range, they decided to focus on an exposed formation of rocks in the Canadian Arctic which were of that age, and which had been deposited in a shallow freshwater environment. The team kept digging year after year. In 2004 they were rewarded with the discovery of remains of a fish they named Tiktaalik. While Tiktaalik is firmly on the fish side of the fish-tetrapod divide, it has an impressive suite of tetrapod-like features.
This was a dramatic vindication of evolutionary theory. A key test of any scientific theory is the ability to make novel predictions which can be experimentally verified. Operating within the conceptual framework of mainstream geology (an old earth) and biology (common ancestry), Shubin’s team picked one location to excavate, and found the type of fossil (previously unknown) that they had predicted. There is nothing in YE creationism or Intelligent Design which could produce such a precise and novel prediction.
According to Wikipedia, the mixture of both fish and tetrapod characteristics found in Tiktaalik include:
Fish – Fish gills, fish scales, fish fins
“Fishapod” – Half-fish, half-tetrapod limb bones and joints, including something like a wrist joint, and radiating, fish-like fins instead of toes
– Half-fish, half-tetrapod ear region
Tetrapod – Tetrapod rib bones and lungs
– Tetrapod mobile neck with separate pectoral girdle
In 2014, Shubin and co-workers published details on the pelvic girdle and fins. Again, features intermediate between fishes and tetrapods are evident:
The pelves of Tiktaalik are paired and have broad iliac processes, flat and elongate pubes, and acetabulae that form a deep socket rimmed by a robust lip of bone. The pelvis is greatly enlarged relative to other finned tetrapodomorphs. Despite the enlargement and robusticity of the pelvis of Tiktaalik, it retains primitive features such as the lack of both an attachment for the sacral rib and an ischium.
The figure below from Ahlberg and Clack shows the differences in skeletal and other features among the fossils species discussed above. There is a gradual loss of the gill cover (blue), and a reshaping of the skull.

Source: P. E. Ahlberg and J. A. Clack, Nature 440, 747-749 (2006)
The following figure by Kevin Padian, reproduced by Anastasia Thanukos, shows more details in the fins and limbs of these transitional species. A progression is seen from 8 to 7 to 6 digits, as intermediates between fish fins and the 5 digits found in living tetrapods. No particular timescale is given for the ghost lineages that predate the fossil appearance of these species.

Source: K. Padian, Integra Comp Biol 2008; 48: 175-88, reproduced in A. Thanukos, Evolution: Education and Outreach 2009; 2: 84-89.
Here is another figure illustrating fish-tetrapod intermediate forms, which calls out the specific changes in limb structure between the fossils.
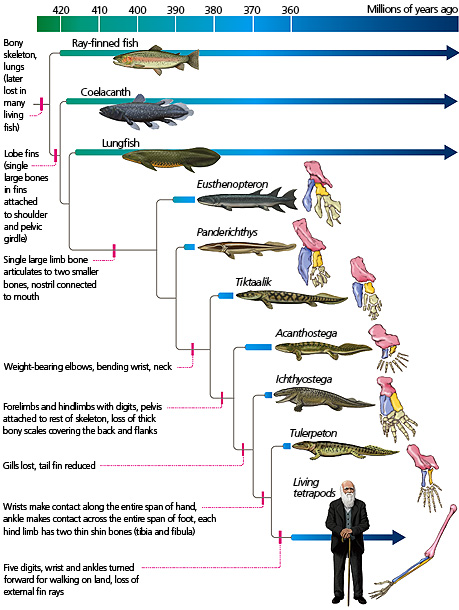
Changes in limb structure among tetrapod fossil intermediates. Source: https://evolution.berkeley.edu/evolibrary/article/evograms_04
The discovery of Tiktaalik does merit accolade, but there is a danger of overstating its role. Some science popularizers have referred to this fish as our “ancestor.” It bears repeating that all the species shown here are dead-end side-branches of evolutionary lineage. As indicated by a careful examination of the figures above, none of them is directly ancestral to living tetrapods. What they do show is that a variety of fishes with some tetrapod characteristics, and a number of early tetrapods with some fish-like features, were present in the geologic strata that predate the appearance of the fossils of modern-type tetrapods. This amount of transition fossil evidence is about what we would expect here, if tetrapods actually did evolve from lobe-finned fishes. This era was over 350 million years ago, which is a lot of time for continents to rise, fall, split, and crash, all of which could destroy early fossils or make them inaccessible.
To bring the tetrapod story up to date, in 2010 Niedźwiedzki et al. published on the discovery of what appear to be tracks of an unknown tetrapod in rocks in Poland that are dated to 397 million years ago. This is about 30 million years before the earliest known primitive tetrapod fossils such as Acanthostega and Ichthyostega. Possible tetrapod tracks, dated to 385 million years ago, have also been found on Valentia Island, off the coast of Ireland. Debate remains over whether these trackways were made by genuine tetrapods. If they were, that pushes the transition from fish to tetrapods some 30 million years earlier than previously believed. YE creationists have claimed that this is a problem for evolution, but it is not. It merely extends the ranges of Tiktaalik, Acanthostega, etc. somewhat further than originally thought.
Friedman and Brazeau have analyzed the patterns of tetrapod-related Devonian fossils, and find evidence that only a small fraction of those species that existed are represented in the known fossils. The preservation rate of species there is about an order of magnitude sparser than for, say, Cenozoic mammals in North America. Thus, it is “unsurprising” that there are “outstanding stratigraphic gaps in the Devonian tetrapod record.”
Below is shown a phylogram from Niedźwiedzki et al. of selected lobe-finned fishes and stem tetrapods, which incorporates these trackway findings. Here Pan= Panderichthys, Tik=Tiktaalik, Aca= Acanthostega, Ich= Ichthyosteg, and Tul = Tulerpeton.

Source: G. Niedźwiedzki et al., Nature 463, 43-48 (2010)
Another possibility is that at least some of the early Devonian trackways were made, not by tetrapods walking on (almost) dry land, but by fish scooting along on their fins in shallow water. King, et al. in 2011 published a study of how some modern lungfish can use their pelvic fins in an alternating pattern to “walk” on the sea floor, with the whole rest of their body held off the bottom. Polish trackway PGI 1728.II.15 (Figure 2 c.) in the Niedźwiedzki paper looks like what a pair of fins might do – – it seems that this trackway was made by a single pair of appendages (not four limbs), with the two appendages sometimes alternating and sometimes pushing along in parallel.
Reptiles to Mammals
The reptile-to-mammal evolution has an interesting twist. There are striking differences between the jaw and ear bone structures of reptiles and of mammals. Reptiles have at least four bones in the lower jaw , while mammals have only one; but reptiles have only one middle ear bone, while mammals have three (the hammer, anvil, and stapes). Studies of developing embryos indicated that two of the bones which, in a reptile get incorporated into a reptilian jaw, in a mammal become part of the middle ear.

Source: Douglas Theobald, “29+ Evidences for Macroevolution”, http://www.TalkOrigins.org; Figure based on K. V. Kardong, Vertebrates: Comparative Anatomy, Function, Evolution. New York: McGraw Hill (2002).
This had scientists in the early twentieth century wondering what kind of evolutionary intermediate could possibly have maintained adequate hearing and jaw function as these two bones gradually moved from the jaw (in reptiles) to the ear (in mammals). The full story is found in the magisterial “29+ Evidences for Macroevolution” article by Douglas Theobald at TalkOrigins.
In the course of the twentieth century a range of fossils were found which documented many stages of the migration of these bones. At one point, there were actually two independent pairs of jaw joints. This was an essential feature that allowed jaw movement while these bones were in intermediate positions.
On the right is one diagram from the TalkOrigins article which illustrates this transition. It shows a side view of three idealized skulls of mammals, therapsids (mammal-like reptiles), and pelycosaurs (early reptiles). The migration of the bones coded purple, yellow, and blue are clear. The location of the jaw joints is shown as black dots. For the therapsid intermediate form, there are two jointed bone interfaces for the jaw.
Many, many fossil intermediates have been found for the reptile-to-mammal transition. According to Kathleen Hunt, ” This is the best-documented transition between vertebrate classes. So far this series is known only as a series of genera or families; the transitions from species to species are not known. But the family sequence is quite complete. Each group is clearly related to both the group that came before, and the group that came after, and yet the sequence is so long that the fossils at the end are astoundingly different from those at the beginning. ” She describes over twenty species that are involved in this transition.
Do Transitional Fossils Prove Evolution?
We have shown for a number of cases, that the fossil record is consistent with evolution, given the four factors (sparse fossil preservation, speciation in small populations, etc.) discussed above. The little changes from one species to the next are not accessible, and direct lineal ancestors are typically not found. However, for a typical major transition a range of “cousin” fossils are found which manifest key intermediate characteristics, in the appropriate time range for that transition. For most objective observers, this constitutes strong supporting evidence for evolution. It certainly shows that the fossil record is not a “problem” for evolution.
What do YE creationists make of these facts? To grind through all the claims and debunking here would take a whole book, so I will just note a few issues. One YE creationist response is to put forth some “fact” which seems to disprove the evolutionary case. Since the horse evolution is so clearly evidenced in the fossils, this seems to call for extra effort by the anti-evolutionists. One such “fact” which has appeared on dozens of YE creationist sites is that the fossils of Hyracotherium (“Eohippus”) are not from an ancient horse ancestor, but are merely the bones of a form of the modern “hyrax” (which lives in the Middle East and looks like a rabbit). This claim is debunked on TalkOrigins here.
Another such “fact” promulgated by YE creationists is that fossils of Hyracotherium and modern Equus have been discovered in the same rock layer. This also is false.
Over the course of the last 150 years of fossil collection and classification, it is inevitable that some mistakes would be made by scientists along the way. For instance, a hundred years ago the figures in most science textbooks showed a single, clean, uniform lineage from little four-toed Hyracotherium to big, one-toed Equus. As noted above, the full picture is more complex than that, with various branching, dead-end lineages, some of which co-existed with each other. Various scientists since 1950 have acknowledged that the earlier simplistic picture of horse evolution was not accurate. YE creationists have taken these statements out of context to make it seem like even “evolutionists” admit that the fossil record does not support evolution.
Deceitfully quoting real scientists is common among YE creationist writers. This “quote mining” enterprise takes statements out of context, and (often making generous use of ellipses) carefully carves out just the words that give the desired effect. There is a large section ( the “Quote Mine Project”) of the TalkOrigins site dedicated to exposing these misleading quotations.
That said, let us circle back to such a quote that we noted at the beginning of this essay: “”The number of intermediate varieties, which have formerly existed must be truly enormous. Why then is not every geological formation and every stratum full of such intermediate links? Geology assuredly does not reveal any such finely graduated organic chain; and this, perhaps is the most obvious and serious objection which can be urged against the theory.”
This quote from Darwin’s Origin of Species is presented endlessly on YE creationist web sites as an admission by Darwin that the facts were against his theory. What the creationists typically fail to include is the very next sentence, in which Darwin tells why this is NOT a problem for his theory: “The explanation lies, as I believe, in the extreme imperfection of the geological record.” This is exactly correct, as we have explained above.
In the next paragraph after these words, Darwin displays a subtle grasp of the nature of likely fossil lineages:
In the first place, it should always be borne in mind what sort of intermediate forms must, on the theory, have formerly existed. I have found it difficult, when looking at any two species, to avoid picturing to myself forms DIRECTLY intermediate between them. But this is a wholly false view; we should always look for forms intermediate between each species and a common but unknown progenitor. [emphasis in the original]
This “common but unknown progenitor” corresponds to the lineal common ancestors which, as noted above, are unlikely to be found as fossils.
These quotes of Darwin occur in the tenth chapter of The Origin of Species, which is appropriately titled “On The Imperfection Of The Geological Record” . Darwin concluded this chapter: “I look at the geological record as a history of the world imperfectly kept and written in a changing dialect. Of this history we possess the last volume alone, relating only to two or three countries. Of this volume, only here and there a short chapter has been preserved, and of each page, only here and there a few lines. Each word of the slowly-changing language, more or less different in the successive chapters, may represent the forms of life, which are entombed in our consecutive formations, and which falsely appear to have been abruptly introduced. On this view the difficulties above discussed are greatly diminished or even disappear.” Right again.
Darwin acknowledged that there were not millions of finely-graded intermediate fossils lying everywhere, yet still a reasonable number of significant transitional fossils had been found. He wrote in Chapter 11 of the 6th edition:
As the accumulation of each formation has often been interrupted, and as long blank intervals have intervened between successive formations, we ought not to expect to find, as I attempted to show in the last chapter, in any one or in any two formations, all the intermediate varieties between the species which appeared at the commencement and close of these periods: but we ought to find after intervals, very long as measured by years, but only moderately long as measured geologically, closely allied forms, or, as they have been called by some authors, representative species; and these assuredly we do find. We find, in short, such evidence of the slow and scarcely sensible mutations of specific forms, as we have the right to expect.
… I have attempted to show that the geological record is extremely imperfect; … that only certain classes of organic beings have been largely preserved in a fossil state; … that, owing to subsidence being almost necessary for the accumulation of deposits rich in fossil species of many kinds, and thick enough to outlast future degradation, great intervals of time must have elapsed between most of our successive formations; …that varieties have at first been local; and lastly, although each species must have passed through numerous transitional stages, it is probable that the periods, during which each underwent modification, though many and long as measured by years, have been short in comparison with the periods during which each remained in an unchanged condition. These causes, taken conjointly, will to a large extent explain why—though we do find many links—we do not find interminable varieties, connecting together all extinct and existing forms by the finest graduated steps. [emphases added]
These citations show that sound reasons for the relative paucity of intermediate fossils (i.e. the inherent sparseness of the fossil record, the likelihood that evolutionary changes mainly take place in small local populations, branching lineages, etc.) were clearly stated in The Origin of Species. My article here has merely fleshed out what Darwin wrote more than 150 years ago. There is therefore no excuse for opponents of evolution who assert or imply that Darwin was acknowledging an actual weakness in his theory.
Varieties of Skepticism
The fossil record is clearly compatible with evolution. Indeed, the same can be said for all physical observations, in geology, biology, chemistry, and genetics. The hard reality, though, is that a dedicated YE creationist will not be convinced of macroevolution by any physical evidence. No matter how many fossil intermediates he is presented with, he will always find a way to wiggle out. He can say, “Yes, there is a sequence of fossils with gradually varying characteristics, but you can’t prove that one evolved into the next; you are just assuming evolutionary relationships among them.” Or, “How do you know that God did not miraculously create these species in this sequence?”
I had some critical things to say about YE creationist Ken Ham’s presentations during his recent debate with Bill Nye. However, I appreciated Ham’s honesty about the basis of his convictions. When he was asked the question, “What, if anything, would ever change your mind?”, he responded: “Well, the answer to that question is ‘I’m a Christian.’ And as a Christian, I can’t prove it to you but God has definitely shown me very clearly through his word and shown himself in the person of Jesus Christ. The bible is the word of God. I admit that that’s where I start from… And so as far as the word of God is concerned, no, no one’s ever gonna convince me that the word of God is not true.” This exemplifies the attitude which renders YE creationists largely impervious to the evidence.
Most Intelligent Design (ID) advocates are motivated by the same sort of conservative Christian faith as YE creationists, but they are not as forthright about their methodology. They claim to be “detecting design”, driven solely by the scientific evidence, but in practice Intelligent Design reduces to the classic god-of-the-gaps. Many Christians find it heartening to learn of various gaps in our current understanding of evolutionary history, as though that somehow demonstrates the necessity of God. Unfortunately, ID consistently overstates the magnitude of these knowledge gaps, relying on deceptive half-truths (e.g. with chimp and gorilla genomes, “junk” DNA, and the Cambrian explosion) to make its case.
ID proponents generally accept the mainstream geological dating of the fossils, which puts them in a quandary: if all those intermediate horse fossils between 5 and 55 million years ago did not result from evolution, how did they get there? If all those reptile-to-mammal transitional fossils back in the Mesozoic do not represent natural evolutionary development, how did they get there? It seems that the Intelligent Agent must have poofed all these species into existence (in a sequence that would later deceive us humans into thinking they had evolved), or else the Agent reached in and physically tinkered with their DNA to make it mutate in unnatural ways. YE creationists may be OK with poofing, but ID advocates are stuck. Since they try to maintain a scientific façade, (ironically) they cannot credit their Intelligent Agent with any actual, specific agency.
Most authors at major YE creationist organizations have some depth exposure to the data of geology and genetics and paleontology. They must actively engage in deception in order to portray these data as compatible with a young earth and incompatible with naturalistic evolution (e.g. see here on Grand Canyon geology).
However, the bulk of the 46% (per recent Gallup poll) of Americans who believe that humans were specially created by God a few thousand years ago are simply going along with their traditional beliefs, reinforced by what they hear from the YE creationist mouthpieces. Furthermore, the evidence for evolution is not as obvious to laymen as they would like to see. We scientists can point to lots of circumstantial evidence, where evolution fits the observed patterns. However, we cannot show a new phylum evolving before their eyes, or even in the course of a 50-year experiment. Nature just does not provide that satisfying level of demonstration of the ability of unguided evolution to cause huge but viable changes in living things in a short time-frame.
What the layman sees with his eyes are animals reproducing “after their kind”, with only minor changes. Scientists can, of course, offer valid reasons for this state of affairs (“Evolution happens to be a really slow process”; “The fossil record is inherently sparse”), but these can come across as lame excuses to someone who is predisposed to doubt evolution.
For perspective, my wife (who is well-educated, but not as a scientist) has commented that, to her anyway, the evidence for evolution seems similar to the nature of the evidence for the Christian faith. By that she meant that there are sound documentary grounds (e.g. some of the letters of Paul) to believe that within a decade of the purported resurrection of Jesus there were people going around risking their lives proclaiming that they had encountered the risen Christ; we have a universe that came into existence at a certain point in time; millions of people today testify to a life-enhancing experience of God; there are videos of extraordinary healings in answer to prayers in the name of Jesus (some of which I have described here and here). These factors are all consistent with the gospel, but do not constitute mathematical proof. (I won’t delve into why God may have set things up such that people must come to Him freely, as opposed to being bludgeoned into faith by frequent miracles or apparitions).
I have read of atheists who sweep aside all the circumstantial evidence that exists in favor of God, and demand to see some miracle like an amputated limb growing back before their eyes. This is unreasonable, at least in relation to the Christian God, since Christ clearly stated that on-demand signs will not be given to unbelievers (Matthew 12:38-39). It is thus similar to the demand of YE creationists to see some spectacular demonstration of accelerated macroevolution. (I suspect that, if these demands were actually met, both the hardcore atheist and the hardcore YE creationist would find some way to discount their experience so as to maintain their previous positions.) My point here is not to justify YE intransigence, but to note that all of us to some degree fit the facts in around our existing opinions.
Addressing the Underlying Issues
A key error of YE creationists is to hold that their interpretation of the Bible is the only valid one. They claim that the only faithful reading of the Genesis creation account is a wooden literalism, and that the six “days” of creation in Genesis 1 were necessarily 24-hour solar days. That is simply not true.
The 24-hour day interpretation was represented among the early church fathers, but it was not the only one. Cyprian (c. 250 A.D.) saw these days as 1000-year ages, Origen (c. 225 A.D.) saw them as figurative, and Augustine sagaciously averred that nobody knows for sure: “What kind of days these were is extremely difficult or perhaps impossible for us to conceive” (City of God 11:6, c. 419 A.D.).
A broad swath of contemporary evangelical Christians (especially in the U.K.), as well as many Orthodox believers and a billion Roman Catholics, hold that a recent, six 24-hour-day creation is not the only faithful Christian view. Evangelist Billy Graham has no problem with God using evolution to accomplish His creative purposes for mankind:
I think that we have made a mistake by thinking the Bible is a scientific book. The Bible is not a book of science. The Bible is a book of Redemption, and of course I accept the Creation story. I believe that God did create the universe. I believe that God created man, and whether it came by an evolutionary process and at a certain point He took this person or being and made him a living soul or not, does not change the fact that God did create man. … whichever way God did it makes no difference as to what man is and man’s relationship to God.
For YE creationists to let go of their false beliefs on the age of the earth, they must either (a) renounce their Christian faith altogether, or (b) find a way to reconcile core Christian beliefs (e.g. those asserted in the Nicene Creed) with the facts of science. Many young adults do reject their faith upon exposure to the evidence of geology and biology, after having been told by parents and pastors (and Ken Ham) that evolution and an old earth are flatly incompatible with the Bible.
Option (b) is, of course, less disruptive. If one is willing to acknowledge that the Bible regularly makes use of metaphor, story-telling, and other figurative language (e.g. Jesus’ parables and Nathan’s confrontation with David), that opens up various options for viewing the Genesis story as meaningful without being literal history. I have outlined here the understanding of Scripture that I have come to, along with links to related resources.
While most YE creationists are driven by their peculiar Bible literalism, the key motivation for the Intelligent Design folks at the Design Institute is more cultural or philosophical. In their foundational “Wedge” document they spell out their dismay over “materialism”, and blame Darwin (among others) for a range of modern problems, up to and including the abuses of communism:
…thinkers such as Charles Darwin, Karl Marx, and Sigmund Freud portrayed humans not as moral and spiritual beings, but as animals or machines who inhabited a universe ruled by purely impersonal forces and whose behavior and very thoughts were dictated by the unbending forces of biology, chemistry, and environment. This materialistic conception of reality eventually infected virtually every area of our culture, from politics and economics to literature and art…. Materialists denied the existence of objective moral standards, claiming that environment dictates our behavior and beliefs. …Materialists also undermined personal responsibility by asserting that human thoughts and behaviors are dictated by our biology and environment. … In the materialist scheme of things, everyone is a victim and no one can be held accountable for his or her actions….Finally, materialism spawned a virulent strain of utopianism. Thinking they could engineer the perfect society through the application of scientific knowledge, materialist reformers advocated coercive government programs that falsely promised to create heaven on earth.
There may be some valid points here, but attacking evolution is not going to fix the problem (“materialism”) that the Design Institute is worried about. Here is why: Every human alive today was conceived and grown by strictly materialistic processes. We can watch sperms fertilize eggs and see the egg cells divide; we can in large measure track the biochemical processes behind all this. The growth of neuronal networks in the fetal brain unfolds without supernatural invention. Every mental event corresponds to some array of physical events in the brain, which in turn are subject to the usual laws of physics. These are the key facts that anti-materialists have to deal with.
Whether or not scientists are able to explain every twist and turn of evolution does not touch these currently-observed facts. Thus, ID’s campaign of sowing doubt about evolution cannot possibly accomplish its avowed goal of unseating materialistic philosophies.
If ID supporters grasped this fundamental point, that could cause the ID movement to refocus in a constructive way. That would be a welcome development, since currently the Intelligent Design movement squanders financial and intellectual resources that might be employed in engaging the real issues with materialism.
******* FOR FIRST-TIME VISITORS TO THIS BLOG *********
Various posts are listed on the right hand side of the main window. Longer essays or letters (e.g. STAN 3) dealing mainly with science/faith issues are accessed by clicking the tabs at the top of the page. Skim the README page to get an overview of what is in these letters. My personal journey of faith-and-science is told here.

Very good work, as usual, and as usual I am going to carp about a minor point that is of interest to me because of my own world view. You state that “I have read of atheists who … demand to see some miracle like an amputated limb growing back before their eyes.” That is an interesting point although stated a bit hyperbolically. I have raised the amputation issue myself, but have not demanded it be performed before my eyes, only that there be creditable evidence of such happening. My point is to ask, if your God can and does perform miracles of healing, why is it that he has never been known to regrow a lost limb? Could it be because unlike some medical problems which our immune systems can handle, regrowth of an amputated limb is not currently medically possible, and difficult to fake creditably?
Most atheists I have read, including Richard Dawkins, are not adamant that nothing could convince them of the existence of a god. The vast majority of us merely claim that the evidence is not there to support the gods who have been proposed. I guess P.Z. Myers might fall under your claim, but I think what he is saying that there is such a massive lack of evidence and implausibility that in an age when David Copperfield can make an elephant disappear on TV (by trickery), there is no single incident which would apply a significant counterweight.
The “we don’t do tests” answer of course is the same answer that cults and their con-artists have long used, so it tends to confirm our suspicions rather than addressing them. As for people being willing to die for mistaken beliefs, and whether that is good evidence for their beliefs, see David Koresh and the Branch Davidians.
I just purchased “How Jesus Became God” by Bart Ehrman. It will present some historical evidence for and against the central premise of Christianity, and it will be interesting to see what conclusions it recommends. (I haven’t read anything specific about its contents so I may be surprised.)
Anyway, I learned something from this post. Maybe some day you will examine historical records of Christianity with the same objectivity and rigor, and the same recognition that people will and have lied for their cherished beliefs. (Or maybe you have, and will post on that.) Thanks for the post.
Jim,
Your comments are thoughtful, as usual. I, too, wrestle with the issue of why there are not more spectacular miracles. For you, I gather it’s an issue of why you don’t see any at all. In my case, I do see occasional notable healings, but wonder why I don’t see more – not in context of proving God’s existence but rather asking whether I should just accept that that is just the way things are or whether there is something I can do to become more effective in praying for others.
So I’ll offer some thoughts on this area, without trying to prove anything. I think it is obvious that if God intended for everyone to be exposed to overwhelming miracles, He could bring that about. But He hasn’t, so either He doesn’t exist (or is otherwise impotent), or has chosen not to for some reason, whether or not we know that reason.
It may be relevant that in the gospels, Jesus does not seem desperate to try to get everyone to follow him. He presented enough teachings to inform those who “had ears to hear”, but many people did not get it. Many of his healings were done because he was “moved with compassion”, not because he was trying to show off. In fact, in some cases he told the healees not to go around talking about it. In John 6, following a crowd-feeding, there was big swell of enthusiasm in the crowd to make Jesus their king (and quartermaster) . Jesus promptly and deliberately offended away nearly everyone by talking about eating his body and drinking his blood, until his followers were whittled down to just a few who valued their connection with him more than having their intellects or sensibilities catered to.
The temptations in the wilderness described at the start of Jesus’ ministry included being challenged to do magical tricks to prove he was the Son of God. Jesus declined. His response was to cite Scripture, instead. When Jesus’ opponents challenged him to do a miracle to prove himself to them, he declined, saying [Mat 12:39] that the most that they would get would be the “sign of Jonah”, i.e. the reports of his resurrection. And he noted in a parable [Luke 16:31] that if someone did not believe the Scriptures, they likely would not believe even if they saw someone raised from the dead.
Things seem to be set up such that there is a decent amount of circumstantial evidence for the gospel (but which, as you point out, can be doubted away if you insist), and no fundamental logical inconsistencies (though plenty of mysteries), yet in the end you have to value God enough to take a step of faith and meet Him on His terms, not yours. And maybe this is not so strange — many of the really important decisions of life are fraught with uncertainty. For instance, it is an act of faith and valuing to freely choose to commit yourself to another human being, for better or for worse, till death do you part.
Jesus [Mat 11:20-24] warned the cities where he had done most of his miracles that, because they had seen these miracles and not repented, their judgment would be more severe. So it may be a form of mercy not to expose people to spectacular miracles that do not change their minds. As the saying goes, be careful what you wish for…
I know that some atheists/agnostics say they are open to change, but as I have noted, it is really, really hard to overcome confirmation bias. Just like YE creationists can always find a way to discount the evidence they are presented with, so atheists can do the same. For instance, I linked in my article above to two earlier posts on videos of healings. The first post ( https://letterstocreationists.wordpress.com/2011/03/10/healings-on-youtube-legs-growing-out/ )
has links to three videos of legs growing out, where we might profitably focus on just the third, short one at the Bethel church in Redding, CA. I also discuss the pitfalls of this particular manifestation — it is very easy to fake or be honestly mistaken. If you want to dig deeper into that last video, here is a more extended version. (Yes, the laughter at the start is kind of grating…)
The camera shot of the key growth spurt is less clear than in the shorter version, but it gives more back story. Here you have a bunch of people who know the girl being prayed for, and know she was originally shorter than the comparison woman, and see by the end she is taller. This is not 100% caught on camera, so you have to decide whether all these people are delusional or in cahoots, or whether the girl really did grow several inches.
My second link ( https://letterstocreationists.wordpress.com/2012/08/14/healing-of-nearly-deaf-boy-on-youtube/ ) goes to a video of an almost-deaf boy in Brazil getting healed. I happen to have known the pray-er, Randy Clark, for many years, and know that he is a man of integrity. The reaction of most atheists to these videos is to not give them a fair viewing, but immediately assert that they *must* be bogus, and then cite extraneous reasons to justify their assertion (e.g. “these things are always fakes” or “extraordinary claims require extraordinary evidence” or whatever). I’m not saying who is right or wrong, just noting the prevailing psychology.
(I should address one other issue: a key reason that you don’t see doctors out there certifying miracles like this is that doctors are very smart people, and they know very well what would happen to them if they did so.)
I can speak from some personal experience that the impact of unusual healings can be less than one might think. Some years ago, I prayed for a young woman who had broken her toe the day before. It was blue and painful. Immediately after the prayer, we watched her toe turn from blue to pink in about 30 seconds, and all pain disappeared. Not the same as a new limb growing, but still pretty impressive. And yet…after that 30 seconds, there was a healthy toe sitting there, and that was that. Unlike in the movies, there were no angelic voices in the background. I was surprised at how unmoving it all was. It did not produce in me a great rush of faith in God, but mainly a sense of, “Well, that was bizarre.”
And (just a few more data points here), despite acknowledging at the time that a remarkable healing had occurred, that young woman drifted away from her Christian faith a few years later, and another young man who witnessed it was not motivated by it to change his unbiblical lifestyle. So this episode left me deeply skeptical as to the evidential value of visible “signs”. I do continue to pray for the sick, but as a means of participating in God’s redemptive purposes, not with the expectation that it will change anyone’s mind.
All this is a long way of saying that (a) there is no reason to believe that the Christian God would or should provide the kind of signs that some people demand, and (b) even if atheists did see signs, it is really unlikely that most of them would actually repent and believe, despite their claims to be open-minded. I’m not insisting I am right here, and there may be exceptions, but that is what my observations suggest.
About Christian sources and Bart Ehrman – – he started off defending fundamentalist Bible literalism, and now he is gotten famous by attacking the same. He presents a lot of interesting stuff, but tends to be ridiculously one-sided.
Yes, there are tens of thousands of textual variations in extant New Testament manuscripts. But – – that is because we have so very many manuscripts spanning many centuries. And the same textual studies that quantify these variations also allow us to determine with a high degree of certainty what the originals were in most cases. Yes, in the process of these textual critical studies we find there are a few beloved passages that we have to discard, and some more that we must amend from traditional translations. But – – there are usually multiple passages that bear on any important point of doctrine, so the net effect of these textual changes is minimal.
Yes, the Gospels look like they were patched together from earlier sources by human writers. But – – that is no shocker, since that’s exactly how Luke 1:1-4 indicates his gospel was written. Yes, the Gospels were written several decades after the events. But — the early letters of Paul (esp. I Cor 15) show that the key aspects of the gospel (atoning death and resurrection of Jesus, the call to believe and follow him in a moral life, offer of forgiveness of sins and hope of resurrection) were well-articulated in the mainstream Christian community within a few years after Jesus’ death.
Yes, the 27 book canon of the New Testament wasn’t officially stated until 327 A.D. But – – the core of this canon, i.e. the Gospels and most letters of Paul, were clearly established by the middle of the second century. We find reverent references to the letters of Paul in the writings of Clement and Ignatius c. 100 A.D., and the four Gospels were so well established by 180 A.D. that Irenaeus referred to the four-fold gospel as being as axiomatic as the four-fold winds of heaven. Whether the last few fringe books like Jude or II John are included is of little import. Yes, there were other “Christianities” competing with the orthodox brand from the first century on. But – – we knew that already, from the most cursory reading of the NT letters and the church fathers.
And so it goes with all of Ehrman’s revelations. They are only problematic if you hold a naive view of inerrancy, are wedded to the King James Bible and its Textus Receptus, and are ignorant of the second-century writings that show most of his claims to be overstated. /rant off/
Peace….and I hope you are enjoying the spring weather.
Wow! Andrea loves horses, but this may be more about them that she wants to know! Let’s dicuss over our fossil fish embedded in the chimney here/ Nate
Scott,
The more I read of your articles… the more you convince me that universal common descent theory is a deception that keeps a Christian in bondage. In fact, I have always leaned Old Earth, but in reading your correspondence and book reviews, I am developing a distant sympathy for young earth – being a long shot that we all missed.
It is sort of like reading Bart Ehrman. I see the other side of your arguments and it is very concerning. When I read a book by Ehrman – I see answer after answer that is no where to be found in the book. When I read your scientific material… I see counter response after counter response that you do not appear to be willing to admit to…
Have you ever seriously fasted and prayed to God that IF universal common descent theory was a “lie” (a very complicated lie evolving hundreds of thousands of comparative inductions), that God would deliver you from this lie?
I ask this sincerely,
Michael
Michael,
I don’t know if you have read “My Story” (Parts 1 and 2) here on my blog, but I can assure you that my engagement in this arena over the past decade has been soaked in prayer. It was with great reluctance that I turned away from the anti-evolution position, knowing that this change would be seen by dear friends and family members as a sort of betrayal. But ultimately I decided that, since God is about truth, I would best honor Him by being rigorously honest about the physical facts as I became more knowledgeable about them.
And it turns out that there is no need to try to defend a literal interpretation of Genesis 1-3 after all – – in the New Testament God the Son communicated primarily by telling stories that did not actually happen, so it should be no surprise that God the Father did so on various occasions in the Old Testament. I celebrate the creation story He gave to the ancient Hebrews as masterfully serving its purpose – – which was to communicate about God and His relation to humans, not to educate us in geology.
I have tried to present the evidence regarding evolution in a fair manner, not trying to oversell it. I made it clear that it is not dramatic as some layman would wish to see, so I can understand why many, perhaps including you, remain skeptical. But evolution has a strong track record in making specific, novel predictions which are then confirmed. This shows its legitimate power.
As for a young earth, that view is simply delusional. If you can’t see that, then you may be in the grip of Morton’s demon (see here: https://letterstocreationists.wordpress.com/grand-canyon-creation/ ), such that prayer and fasting might be useful. Sorry to be so blunt, but I have spent too much time on the geological issues to pretend that they are ambiguous. Of course, if you have discovered new evidence that the earth is young, the news media would be open to airing it. I noticed that TIME published an op-ed piece by Ken Ham on the recent Noah movie.
As you are sorting these things out, I commend to you the observation of Saint Augustine on the collateral damage done when well-meaning Christians feel they must defend a literal view of Scripture in opposition to the established body of physical knowledge:
“ Usually, even a non-Christian knows something about the Earth, the Heavens, and the other elements of this world, about the motion and orbit of the stars and even their size and relative positions, about the predictable eclipses of the sun and moon, the cycles of the years and the seasons, about the kinds of animals, shrubs, stones, and so forth, and this knowledge he holds to as being certain from reason and experience. Now, it is a disgraceful and dangerous thing for an infidel to hear a Christian, presumably giving the meaning of Holy Scripture, talking nonsense on these topics; and we should take all means to prevent such an embarrassing situation, in which people show up vast ignorance in a Christian and laugh it to scorn.
“ The shame is not so much that an ignorant individual is derided, but that people outside the household of the faith think our sacred writers held such opinions, and, to the great loss of those for whose salvation we toil, the writers of our Scripture are criticized and rejected as unlearned men. If they find a Christian mistaken in a field which they themselves know well and hear him maintaining his foolish opinions about our books, how are they going to believe those books in matters concerning the resurrection of the dead, the hope of eternal life, and the kingdom of heaven, when they think their pages are full of falsehoods on facts which they themselves have learnt from experience and the light of reason?” – St. Augustine, The Literal Meaning of Genesis (408 A.D) Book 1, ch.19.
May the Lord grant you peaceable wisdom in these matters…
Scott,
thank you for answering.
I love reading your material, and also Gerard Jellison’s book reviews.
I know that you both believe you are doing a service to set the record straight regarding creationism, I primarily only read anti-creationist pro-evolutionary material.
I love you guys and I pray for you both. I see the complications differently than you guys. I also see the whole history of science differently than you do. If we both love Jesus than we can be unified in Him. You and I could debate 10,000 specifics in all of the various fields of research but we both have different roles right now in the scope of God’s perfect plan to work through (in spite of) our imperfections.
I could accuse you of being systematically deceived by tens of thousands of inductions which are open to error and you could accuse me of being ignorant of science, not knowing what I’ve learned in recent years…. or how I process the same information differently.
Just out of curiosity, what if someone like Francis Collins converted to creationism and decided to agree with the Sanford’s and the Meyers of the world? How would you respond to someone like Francis Collins IF he made that type of accusation of deception?
What do you think of when someone claims you are deceived because of naturalistic assumptions which were influence all of your scientific educational upbringing?
In Him,
Michael
Michael, [reply to your April 17 comment]
As noted previously, essentially all physical observations of rocks, fossils in the rocks, light from distant stars, and genomes accord with the old earth/evolution viewpoint. I say “essentially all” because at any point in time there is a “bleeding edge” of our expanding envelope of knowledge where there are observed facts which are not yet understood in terms of the greater whole of the uniform processes of the space-time continuum.. The trend is that, with time, these knowledge gaps get filled in, all in terms of naturalistic processes. For instance, in the article above, there was a time when it seemed that evolution was in trouble as far as the reptile-to-mammal transition. How could you possibly get two bones to migrate from the jaw in reptiles to the inner ear in mammals, and still have a functional jaw or ear?
YE creationists love to play up the signficance of today’s knowledge gaps, as though these gaps are some sort of support for young earth/anti-evolutionism. There are dozens of such items they have claimed in the past (amount of salt in the ocean, fluctuations in the earth’s magnetic field, distance from the moon to the earth, etc. etc.), and some they continue to claim. Nearly all of these gaps or problems of yesteryear have now been explained within the framework of science. YE creationists differ in their level of honesty in admitting that. I have detailed their rank dishonesty regarding Grand Canyon geology, as an example . At the other end of the honesty spectrum in YE creationism stands Kurt Wise, a bona fide PhD (Harvard) geologist, who on the one hand acknowledges “Substantial supporting evidence of macroevolutionary theory can be found in the fossil record of stratomorphic intermediates” (i.e. he does not try to fib away what I presented above), yet makes it clear that no amount of physical evidence for evolution would ever make him believe in it: “If all the evidence in the universe turns against creationism, I would be the first to admit it, but I would still be a creationist because that is what the Word of God seems to indicate. Here I must stand.”
To repeat: essentially all the physical evidence comports with an old earth and evolution, within an intellectual framework which assumes that the same laws of physics we observe today have held in the past. The light from distant stars, and the rock layers, serve as time machines to test that assumption of the invariance of physics, and indeed it holds, at least for the past 13 billion years.
What you call “naturalistic assumptions” is simply that intellectual framework, namely that there is consistency in the natural world. I would remind you that this framework, in its modern scientific form, was not a pagan invention, but developed out of Christian thought in Europe several centuries ago (see, e.g. Francis Bacon, Kepler, etc). Indeed, the only way that we can have miracles that operate as signs of God’s special intervention is for the norm to be the monotonous regularity of physical operations. And that is clearly the New Testament framework: Jesus said, “Do not believe me unless I do the works of my Father. But if I do them, even though you do not believe me, believe the works, that you may know and understand that the Father is in me, and I in the Father.” (John 10:37-38). And when Paul was trying to convince the Corinthians (I Cor 15) of the reality of the resurrection, he did not say, “Just believe me because my opinion is correct,” but rather he cited the testimony of eyewitnesses, some of whom were still alive at the time.
If you simply refuse to believe that the overall conclusions of modern science are real, despite the fact that “tens of thousands” of observations are readily explained within that framework (and not in YE/Flood geology), AND it makes bold predictions which are then confirmed, then I don’t know what to say. There are people who refuse to believe that the Apollo moon landings really happened — they find ways to dismiss all the videos and testimonies and moon rocks as mere circumstantial evidence that could be interpreted differently. Fine. There are all kinds of levels of skepticism that one can indulge in — how do I know that the outside world really exists at all ? Maybe it is just a figment of my imagination…. But these sorts of head games are not biblical: the New Testament view of the physical world is a common-sense, realistic one, else John’s insistence that Jesus came in the flesh would be meaningless blather.
The only honest alternative that I can see to theistic evolution is the Appearance of Age position, which admits that the universe looks old (including light arriving from supernovae events millions of years ago) and that it looks like we got here by evolution (with beat-up looking genomes), and graciously admits to the predictive power of modern science, yet holds that this all is an illusion — God created the universe in 4000 B.C. with all these old-looking physical features, and after the global Flood, God rejiggered human genomes to make it look like the Ark genetic bottleneck (the whole earth repopulated from the six younger folks on the ark c. 2500 B.C.) never happened.
But this campaign of divine illusion seems to me less in keeping with God’s wise and truthful character than the notion that God, in the Genesis story, gave the ancient Hebrews a parable that met their needs at the time, and that future generations of His people were supposed to use their God-given minds to figure that out and applaud it instead of denounce it.
re:”How would you respond to someone like Francis Collins IF he made that type of accusation of deception?” – – Since childhood I have been an independent thinker, evaluating things for myself, so I would not be particularly impressed if any particular person swung one way or the other.
I hope this answers your questions.
Best regards,
Scott
Bondage to reality?
BTW, I haven’t seen it in your archives yet, but I was wondering if you ever critiqued this article years ago? http://www.answersingenesis.org/articles/tj/v14/n3/pseudogenes-genomes
Michael,
No, I have not addressed that article, and do not intend to, for several reasons. First, it was written in 2000, well before the full sequencing of genomes was available, so it would be superceded by better data by now. Second, it is authored by the pseudonymous “John Woodmorappe”, who is actually Illinois high school teacher Jan Peczkis, who has a dismal record misrepresenting the evidence in a field I know more about (namely, geology). A fair assessment of his geology was stated as: “Many of his arguments contain quotes out of context, scientific data out of context, and omissions of key points which would invalidate his arguments.” (http://www.oldearth.org/profiles/john_woodmorappe.htm ). This doesn’t necessarily mean he was wrong also in this 2000 article, but it makes me less motivated to dig in to find out.
The main reason, though, is that I have already spent hundreds of hours investigating, in depth, a number of anti-evolution claims. These include gorilla/chimp/human genome comparisons, Cambrian fossils, “junk” DNA, and the complex of assertions in Genetic Entropy. In every case I have found their claims to be false, being based entirely on partial truths stemming from ignorance or outright deceit. Ditto for investigating many young earth geology claims. Having gone through this experience again and again, it is hard to get excited over some other “betcha can’t explain THIS” article by the anti-evolutionists.
Again, if you have come across genuine evidence that disproves evolution, feel free to share it with the world. Fame awaits you. If not, you might think seriously about re-examining your approach to the Bible in the light of what II Tim 3:15-17 states as its purpose.
Wishing you well,
Scott
P.S. You might enjoy the movie “God’s Not Dead” – – it deals, among other things, with the implications of the Big Bang. The overall plot has to do with a Christian college freshman who is confronted with an atheistic professor who requests that all the students in his philosophy class sign to the statement that “God is dead”, but it has a lot of other subplots that deal with the realities of aging, life choices, and facing one’s own mortality.
From those philosophically minded that I know whom have seen it, it does not present a comprehensive theodicy according to them. I haven’t seen it yet.
BTW, I wanted an honest rebuttal to the AIG link. It was not a “betcha can’t explain THIS article” nonsense. I don’t want to discuss the material until I see it fully responded to.
I will say that I think the “fame awaits you” line demonstrates that you really do not understand what is going on here with differentiating between what looks like science and what really is observable and falsifiable. I also feel you do not seem to be aware of the bias of peer review and other UCD theory advocates/infrastructures which refuse to acknowledge the obvious observations regarding deleterious mutations.
What are your view’s on Marcus Ross PhD in your similar field of study? Have you responded to any of his work or curriculum?
Once again a great article and well written. My church is obsessively OEC and my internal journey has been very painful (and stressful for my dear wife). Alas I think it will get more painful as others react in fear to my quietly held opinions. Thankyou very much for sharing your journey and knowledge. It is a valuable work. Daniel
Daniel,
Well, at least they are not YEC! 🙂
Seriously, I do empathize. I’ve learned to always preface my pro-evolution comments around my conservative friends with some sort of context-setting remarks, like briefly recapping my journey or noting how Paul put a high value on empirical evidence (e.g. I Cor 15). If I don’t have enough time to make a reasonable case, it is not worth saying anything b/c it will just be misunderstood. My wife needed to hear me say over and over again that I placed a high value on the Genesis story, though as a parable rather than as history.
It can be pretty lonely as evangelical Christian who is science-literate. In the U.S. (American Scientific Affliliation), U.K. (Christians in Science), and Canada there are associations of folks like us. I enjoy reading the ASA journal, and gathering with them every other year.
Best wishes…
Scott
Pingback: A Survey of Biblical Natural Theology | Letters to Creationists
if we will find a self replicating watch with dna. is this kind of watch will be evidence for design or evolution?
Pingback: Exposing the Roots of Young Earth Creationism | Letters to Creationists
Pingback: “Big Daddy” Chick Tract: The Most Widely-Distributed Anti-Evolution Publication | Letters to Creationists
Pingback: A Creationist Speaker Comes to Town | Letters to Creationists
Pingback: Listing of Articles on Science, Faith and Other | Letters to Creationists
Pingback: Evolution Before Our Eyes: Complex Mutations in Microbes Giving New Functions | Letters to Creationists
Reblogged this on Quaerere Propter Vērum.
Pingback: Evolution and Faith: My Story, Part 2 | Letters to Creationists
Pingback: Science and Faith at the American Scientific Affiliation 2018 Meeting | Letters to Creationists
Pingback: Tanis Site: The Day the Dinosaur-Killing Asteroid Hit | Letters to Creationists
Pingback: 2019 Letters to a Creationist, Part 2 | Letters to Creationists
Pingback: How to challenge a scientific theory, method 1: Evidence that contradicts it – How old is the earth?
Pingback: Saint Augustine on Interpreting Genesis | Letters to Creationists
Pingback: Whale Origins: A Test Case for Evolution | Letters to Creationists
Pingback: Fazale Rana Disputes Evolution [2020 NCCA Apologetics Confc, 3] | Letters to Creationists
Pingback: John Sanford Disputes Evolution [2020 NCCA Apologetics Confc, 5] | Letters to Creationists
Pingback: 低端护教学(6)|启底年轻地球论(二) – Eddy & Emma's Blog