
Cambrian Contention: Disputing “Darwin’s Doubt”
In June, 2013 Stephen Meyer of the Discovery Institute published Darwin’s Doubt. This book reviews some of the facts regarding the relatively sudden appearance in the fossil record of many major groupings (“phyla”) of animals in the Cambrian period. Meyer claims that this pattern cannot be handled within the mainstream theory of evolution, claiming that “Intelligent Design” (ID) provides a better explanation.
The Publication of Darwin’s Doubt
The Discovery Institute (DI) worked hard to promote Darwin’s Doubt. The DI bombarded its supporters with emails urging them to pre-order the book — I know, since I am on one of their email distribution lists. The DI also engaged in a variety of activities to heighten awareness, asking their supporters to tell friends and family about the book and encouraging them to pre-order copies, and also asking for donations to support promotional activities such online advertising, production of podcasts, etc. The claim was that, “With your help, this book will change the course of the origins debate for generations to come.”
This intense promotion of Darwin’s Doubt was successful, at least in the short term. The book was released on June 18, and quickly shot to #7 on the New York Times top 25 best-seller list for hard-cover non-fiction for the week of July 7. Its sales quickly sank after this promotional pop was exhausted. One week later the book had ignominiously disappeared altogether from the NY Times bestseller list, but the DI still touts it as a “New York Times Bestseller”. As of early October, Darwin’s Doubt stood at about #1000 in Amazon sales. It still ranks #1 or # 2 in Amazon’s categories of Theology>>Creationism, and (to the chagrin of practicing scientists) in the categories Evolution>>Organic, and Biological Sciences>>Paleontology.
The contents of Darwin’s Doubt are summarized in this sympathetic review in the American Spectator , and also in the many favorable reviews on Amazon. Here is a very detailed chapter by chapter summary of Darwin’s Doubt, with dozens of excerpts.
To many readers, Meyer makes a convincing case. He notes the first appearance of fossils of many animal classes in the middle of the Cambrian period, in a timeframe of 5-10 million years, which is short by geological standards. In most cases, there are no unambiguous antecedents to these animals in the fossil record. He notes the extreme complexity and interdependence in genetic and epigenetic systems, which make it difficult to make a change in one aspect without degrading the overall functioning. He cites the experimental results of Douglas Axe, which seem to show that it is extremely difficult to move, by small but viable increments, from one form of a gene to significantly different form via random mutations. As Meyer presents it, many major animal groups spring into existence in a time-frame far too short to be accounted for by the standard evolutionary mechanisms of random mutation and natural selection. If Darwinian evolution fails to explain the explosion of new genetic information, Meyer recommends consideration of another, well-proven source of information, which is design by some intelligent agent.
This book was intended to provoke controversy, and in this it has been successful. Strongly-worded critical reviews have been answered by equally pointed essays from supporters of ID, as pro- and anti-evolution forces battle for the hearts and minds of the public.
It is written in the book of Proverbs (18:17), “In a lawsuit the first to speak seems right, until someone comes forward and cross-examines”. In my efforts to dig down to the truths regarding evolution, I have found it useful to follow on-line arguments back and forth. As each side attacks the other and defends itself, it eventually becomes clear what is reality as opposed to rhetoric. In that spirit, I will summarize here some reviews that are critical of Darwin’s Doubt, and also some responses by the book’s defenders. But first, a little technical background.
The Nature of the Fossil Record
It is a general truism that a species appears suddenly in the fossil record without a clear, graduated set of intermediate forms between it and some previous species. This is precisely what we expect from the following two facts:
(1) Very, very, very few of all the organisms that have died in past eons become fossilized. As we can observe today, nearly all carcasses rot or are eaten by scavengers rather than being buried intact in rock layers. Of the remains that do get fossilized, many are later eroded away, or smeared beyond recognition in metamorphic transformations deep in the earth. This is more of an issue for older rock layers, such as the Cambrian and Precambrian, since they have had more time to be either raised up to the surface for erosion or to be buried more deeply under other rocks. Also, of all the potential fossil-bearing rocks, only a small fraction of them are available near the surface for paleontologists to examine.
The Coelacanth order of fishes furnishes a classic example of the fickleness of the fossil record. The Coelacanth order of fishes was once widespread in the ancient seas. Coelacanths peaked in the fossil record about 240 million years ago, and then declined. The most recent known fossil dates back to about 80 million years ago. It was thought that they had become extinct. In 1938, however, a live coelacanth was discovered in the Indian Ocean. Since then a number of others have been caught. Unless we are prepared to claim that an Intelligent Agent supernaturally re-created these modern coelacanths, we must acknowledge that some population of these fish has existed for the past 80 million years but without leaving a trace in the fossil record.
(2) Basic population genetics shows that it is difficult for new genetic mutations to become established in very large populations. Thus, it is far more likely that a new species would develop within a small, isolated population, especially if that population is under some environmental stress that would favor genetic changes. If the new species becomes more fit than the old species, the new species will expand and only then is likely to appear in the fossil record. But once a species is widespread and successful in its ecological niche, there will be diminished selection pressure for changes, so fossils of this now well-adapted species are likely to appear for perhaps million of years with showing little change.
Evolutionary lineages tend to be “branchy”. Typically the organisms on the side branches show up in the fossil record, rather than the transitional ones along the main “trunk”.
Thus, we should expect lots of gaps in the known fossil record. The fact that various transitional fossils have not been found is not a rational basis for believing that these transitional forms never existed.
The fossils that HAVE been found fit well within this conceptual framework. There is a clear, general progression of life-forms found in the fossil record. Trilobites (early arthropods, distant cousins to today’s insects and crustaceans) always appear in sedimentary layers below any rocks containing dinosaurs (reptiles), never above. In the vertebrate lineage, first fish, then tetrapod amphibians, then reptiles, and finally mammals make their appearance. A number of fossils with part fish/part amphibian characteristics, and with part reptile/part mammal characteristics, are found at about the times expected for these transitions. This is all consistent with common ancestry within the theory of evolution.
When we zoom into the details the appearance of individual species, however, the fossil record appears “jumpy” rather than smoothly continuous. Anti-evolutionists claim that this disproves evolution. However, as explained above, this pattern is consistent with the facts that (1) the fossil record is known to be extremely incomplete, and (2) transitional forms are most likely to arise in small populations. Thus, it is quite reasonable to hold that transitional species did exist, but not in sufficient amounts or locales to leave discoverable fossils. The alternative to evolution is that some Intelligent Agent created, and then quickly killed off, species after species after species over hundreds of millions of years, where the morphological sequences of these species just happen (!) to comport with expectations from common descent via evolution (e.g. the observed fish/amphibian/reptile/mammal sequence, as opposed to fossilized rabbits randomly appearing in 500 million-year-old rocks).
The Cambrian Explosion
Geologists have divided the ages of the sedimentary rock layers into a number of divisions (ages, periods, epochs, etc.) that help in their study. These divisions are generally distinguished by the characteristic sets of fossils found in their rocks. The Cambrian period is seen as lasting from about 542 to 485 million years before the present. In this period many of the major groupings (“phyla”) of animals first appeared in the fossil record. This phenomenon is sometimes called the “Cambrian explosion.” The time before the Cambrian, the Precambrian, shows relatively few fossils of animals other than traces of worm-like creatures.
When Darwin published The Origin of Species in 1859, almost no animal fossils were known from the Precambrian. He acknowledged that the relatively sudden appearance of large, complex life-forms in the Cambrian (then considered part of the “Silurian” period) posed a challenge to his theory of evolution. He attributed the general lack of transitional forms to the inherent incompleteness of the fossil record. For the specific case of the Cambrian, Darwin suggested that for some unknown reason most of the rock layers immediately prior to the Cambrian (which presumably contained the expected transitional fossils) were missing. (It turns out that Darwin’s conjecture has been verified – due to continental uplifts, there was massive erosion of previous rocks around the beginning of the Cambrian which would have destroyed a lot of potentially fossil-bearing sedimentary layers, but scientists don’t believe that this is the major explanation for the Cambrian explosion.)
The situation has changed immensely since Darwin’s day. A great number of fossils of Precambrian life-forms have been discovered in the past hundred years. These comport with expectations from the theory of evolution: the earlier fossils represent the simplest organisms, such as single-celled bacteria, starting about 3.4 billion years ago, then more complex cells such as eukaryotes (2 billion years ago), and multi-celled organisms (1 billion years ago). Starting about 600 million years ago, in the Ediacaran period which immediately preceded the Cambrian, reasonably large animals appear.
It has been very difficult for biologists to classify the Ediacaran fauna, since most do not clearly fit within categories of later animals. Most of them vanish from the fossil record with the advent of the Cambrian life-forms. A few representatives of more modern organisms do appear in the Ediacaran. For instance, fossilized sponges, animals representing the modern phylum Porifera, are found in the Ediacaran. Well-preserved fossil sponges from about 580 million years ago in the Ediacaran period have been found in the Doushantuo Formation. Many diversified sponge fossils are found in the early Cambrian. Closer to the vertebrate family line, Kimberella appears in rocks that about 560 million years old, some 20 million years before the start of the Cambrian. Wikipedia has pictures of Kimberella fossil remains and of an artist’s reconstruction of the slug in 3D, reproduced here:

This slug may or may not be a primitive form of mollusk, but it is generally accepted to be a bilaterian. Bilaterians are distinguished by (at at least one point in their life cycle) having left-right symmetry, and distinct front/back and top/bottom. Nearly all the macroscopic animal phyla, including chordates and arthropods, that appear in the Cambrian are bilaterians. They all share a great many genes. While Kimberella may not be directly ancestral to these later animals, its fossils demonstrate that the basic genetic package of bilaterians already existed well before the start of the Cambrian. Thus, the development of the Cambrian life-forms was not a gigantic ab-initio enterprise, but a diversification of already-existing genomes.
The Cambrian explosion has long been a staple of creationist arguments. The relatively sudden appearance in the fossil record of many phyla is presented as a disproof of evolution. It is not. This topic has been treated at length, e.g. in a series on the Biologos web site.
The first appearance of ANY phylum in the fossil record is, superficially, a “challenge” to evolution. It makes little difference, logically, whether the first appearance of various phyla is spread out evenly over the ages or is concentrated in some particular period. As usual, anti-evolutionists are reduced to complaining about the fossils that have not yet been found, while failing to properly evaluate the fossils that have been found. We have explained above why the lack of transitional fossils is not a problem.
Here is a chart from Berkeley of the first appearance of fossil representatives of all the phyla of today’s metazoans (i.e. most animals, apart from protozoa). Cnidaria (corals, jellyfish) appear in the Precambrian. So do Porifera (e.g. sponges), even though they mistakenly appear in this chart at the very start of the Cambrian. Thus, ten of these phyla first appear here within a 50 million year span of the Cambrian, with eight of these ten appearing in a 5-10 million year window about a third of the way through the Cambrian. In addition, fossils in the Cambrian and Precambrian are found which scientists cannot clearly fit into any existing phylum, so they are placed in several other phyla that later go extinct. Throughout the remainder of geological history, another eight phyla appear in the fossil record. Another twelve phyla, at the right edge of the plot, are represented by living species for which no fossils have been discovered. These are mainly small, soft-bodied creatures such as roundworms or flatworms.

Anti-evolutionists cite quotes by scientists on how “amazing” it is that ten major animal phyla appear within the Cambrian period. Or how “astonishing” is the first appearance of eight of those phyla in a concentrated span of 5-10 million years in the middle Cambrian, with perhaps some extra phyla that have since gone extinct. I, too, feel the wonder, but it is the same sort of wonder I felt when I first laid eyes on the Grand Canyon. The Grand Canyon is impressive, surprising to come across, and not entirely explained yet by science (there is disagreement on how old it is , and no one really knows where the Colorado River, if there was one back then, was flowing prior to the uplift of the Colorado Plateau millions of years ago). However, the wonder and the mystery would not justify abandoning a scientific framework for the erosion of the Canyon, and believing that only an alien spaceship or a giant supernatural finger reaching down from heaven could have carved out such a spectacle.
If sheer rapidity of appearance to human observers is a measure of importance, the Cambrian explosion pales in significance compared to the “Holocene Detonation”, i.e. the discovery within the past thousand or so years of the eleven animal phyla which have left no fossil trace. These are the phyla that are clustered along the lower right hand side of the diagram above.
How can we account for the Holocene Detonation? It is of course possible that an Intelligent Agent suddenly created flatworms and roundworms a thousand years ago, and that is why we find no fossils of them. Mainstream science believes that flatworms and roundworms have existed for many millions of years, but simply have not been suitably preserved in sedimentary rocks (see above on the inherent paucity of the fossil record). Similarly, the first appearance of arthropods and other phyla in the middle Cambrian does not mean that no transitional species for these groupings existed before then. As noted above, it is entirely reasonable to assume that these intermediate forms did exist but do not show in fossils.
Geologists have found evidence that at about the beginning of the Cambrian period, there was a marked rise in the level of calcium in the oceans, probably due to plate tectonic events leading to enhanced uplift and erosion of continental rocks. The higher calcium concentrations would make it easier for organisms to form hard shells. Also around this time, the oxygen level in the oceans increased, which would help facilitate the appearance of larger, more active species. These environmental factors, of course, would not directly make genomes mutate and diversify. However, many lines of evidence show the existence of a diverse, mainly worm-like animal community in the late Ediacaran, and larger animals with some shelly parts in the earliest Cambrian. It is reasonable to suppose that some of those worms or slugs had (over preceding millions of years) developed the genetic features characteristic of bilaterian animals (e.g. Pax-6 gene for eyes, hox genes for anterior-posterior body plan), and that over the first 20 million years of the Cambrian these wormish critters diversified and developed harder body parts. Note that for organisms that likely reproduced at least once a year, 20 million years is a lot of generations.
Lagerstätten: Places of Exceptional Soft-Bodied Fossil Preservation
It is crucial to understand that not all sedimentary rock layers are equally good at preserving fossils. As already noted, the usual fate of a carcass is to rot or be eaten. Thus, it is most unusual to find fossilized impressions of soft-bodied animals. There are some rare but important exceptions to this rule: in a few places in the world there are sedimentary deposits which provide exquisite preservation of soft as well as hard body parts. This type of place is called a “Lagerstätte”, which is a German word meaning “storage place” (plural: “Lagerstätten”).
From Wikipedia, “Cambrian Explosion”:
The Cambrian fossil record includes an unusually high number of lagerstätten, which preserve soft tissues. These allow palæontologists to examine the internal anatomy of animals, which in other sediments are only represented by shells, spines, claws, etc. – if they are preserved at all…While lagerstätten preserve far more than the conventional fossil record, they are far from complete. Because lagerstätten are restricted to a narrow range of environments (where soft-bodied organisms can be preserved very quickly, e.g. by mudslides), most animals are probably not represented; further, the exceptional conditions that create lagerstätten probably do not represent normal living conditions.…The sparseness of the fossil record means that organisms usually exist long before they are found in the fossil record – this is known as the Signor–Lipps effect.
The world’s major Precambrian and Cambrian Lagerstätten include:
Precambrian Lagerstätten
Bitter Springs 1000–850 Mya South Australia
Doushantuo Formation 600–555 Mya Guizhou Province, China
Mistaken Point 565 Mya Newfoundland, Canada
Ediacara Hills 550-545? Mya South Australia
Cambrian Lagerstätten
Maotianshan Shales (Chengjiang) 518 Mya Yunnan Prov., China
Sirius Passet 518 Mya Greenland
Emu Bay Shale 517 Mya South Australia
Kaili Formation 513–501 Mya Guizhou prov., south-west China
Blackberry Hill ~510–500 Mya Central Wisconsin, US
Wheeler Shale (House Range) 507 Mya Western Utah, US
Burgess Shale 505 Mya British Columbia, Canada
Kinnekulle Orsten and Alum Shale 500 Mya Sweden
Öland Orste and Alum Shale 500 Mya Sweden
One point to note is that these Lagerstätte are fairly rare. It happens that there is a cluster of them 500-525 million years ago (Mya) in the middle of the Cambrian, but that is an anomaly. Through most of the past 500 million years, we find them maybe once every 15 million years. Another key point is that there is a 20 million year gap between the dates of the late Precambrian Ediacara formation, and the Maotianshan (Chengjiang) sediments. The Chengjiang assemblage represents the key burst of the Cambrian explosion in the fossils; this is where many of the animal phyla first appear. However, this burst is somewhat misleading – it is in part an artifact of the lack, in the 20 million years preceding the Chengjiang, of a sedimentary formation with high-quality preservation characteristics.
Meyer’s treatment of the fossil record is therefore quite inadequate. He claims that, if the predecessors to the mid-Cambrian fauna existed, we should be able to find them in the fossil record. This claim is false. He writes (Chapter 3), “The discoveries near Chengjian demonstrated beyond any reasonable doubt that sedimentary rocks can preserve soft-bodied fossils of great antiquity and in exquisite detail, thereby challenging the idea that the absence of Precambrian ancestors is a consequence of the fossil record’s inability to preserve soft-bodied animals from that period.” This is a misleading half-truth. Yes, soft-bodied animals “can” be preserved in exquisite detail. But this only occurred under very rare circumstances, and it so happens that those circumstances (to our knowledge) did not occur in the crucial first 20 million years of the Cambrian when the predecessors of the Chengjiang fauna were developing. Also, those circumstances occurred only a few times in the entire preceding 500 million years of the Precambrian, so it is unreasonable to expect to find a complete sampling of the Precambrian fauna in the fossil record.
Let the Games Begin
As soon as Darwin’s Doubt was released, supporters (as urged by the Discovery Institute) and critics started posting reviews on Amazon. Most of the early critical reviews focused on issues around the Cambrian fossils, since that was the main novel edge of the book. Less attention was paid to the arguments that Meyer makes against the adequacy of unguided evolution to produce big changes in genomes. Likely this was because these are the same arguments that ID advocates have been making for years, and which have been answered in other venues. For instance, here Steve Matheson politely explains why the Douglas Axe experiments which Meyer cites as demonstrating the near-impossibility of evolving new proteins in fact demonstrate no such thing.
A key critical review on Amazon was posted by geologist and author Donald Prothero, aggressively titled “Stephen Meyer’s Fumbling Bumbling Cambrian Amateur Follies”.
The titles of most reviews, whether attacking or defending the book, are in-your-face; many critical reviews, including this one by Prothero, are laced with venomous insults and accusations against Meyer and other ID proponents. This review, published on July 21, became a lightning rod for supporters and critics of the book, with over 2700 comments posted on it. Prothero’s core critique is this:
The entire diversification of life is now known to have gone through a number of distinct steps, from the first fossils of simple bacterial life 3.5 billion years old, to the first multicellular animals 700 m.y. ago (the Ediacara fauna), to the first evidence of skeletonized fossils (tiny fragments of small shells, nicknamed the “little shellies”) at the beginning of the Cambrian, 545 m.y. ago (the Nemakit-Daldynian and Tommotian stages of the Cambrian), to the third stage of the Cambrian (Atdabanian, 530 m.y. ago), when you find the first fossils of the larger animals with hard shells, such as trilobites. But does Meyer reflect this modern understanding of the subject? No! His figures (e.g., Figs. 2.5, 2.6, 3.8) portray the “explosion” as if it happened all at once, showing that he has paid no attention to the past 70 years of discoveries. He dismisses the Ediacara fauna as not clearly related to living phyla (a point that is still debated among paleontologists), but its very existence is fatal to the creationist falsehood that multicellular animals appeared all at once in the fossil record with no predecessors. Even more damning, Meyer completely ignores the existence of the first two stages of the Cambrian (nowhere are they even mentioned in the book, or the index) and talks about the Atdabanian stage as if it were the entire Cambrian all by itself. His misleading figures (e.g., Fig. 2.5, 2.6, 3.8) imply that there were no modern phyla in existence until the trilobites diversified in the Atdabanian. Sorry, but that’s a flat out lie. Even a casual glance at any modern diagram of life’s diversification (Figure 1) demonstrates that probable arthropods, cnidarians, and echinoderms are present in the Ediacara fauna, mollusks and sponges are well documented from the Nemakit-Daldynian Stage, and brachiopods and archaeocyathids appear in the Tommotian Stage–all millions of years before Meyer’s incorrectly defined “Cambrian explosion” in the Atdabanian. The phyla that he lists in Fig. 2.6 as “explosively” appearing in the Atdabanian stages all actually appeared much earlier–or they are soft-bodied phyla from the Chinese Chengjiang fauna, whose first appearance artificially inflates the count. Meyer deliberately and dishonestly distorts the story by implying that these soft-bodied animals appeared all at once, when he knows that this is an artifact of preservation. It’s just an accident that there are no extraordinary soft-bodied faunas preserved before Chengjiang, so we simply have no fossils demonstrating their true first appearance, which occurred much earlier based on molecular evidence.
Meyer’s distorted and false view of conflating the entire Early Cambrian (545-520 m.y. ago) as consisting of only the third stage of the Early Cambrian (Atdabanian, 530-525 m.y. ago) creates a fundamental lie that falsifies everything else he says in the ensuing chapters.
To help make sense of this, here is a figure the Prothero posted elsewhere which illustrates the time divisions he is talking about:
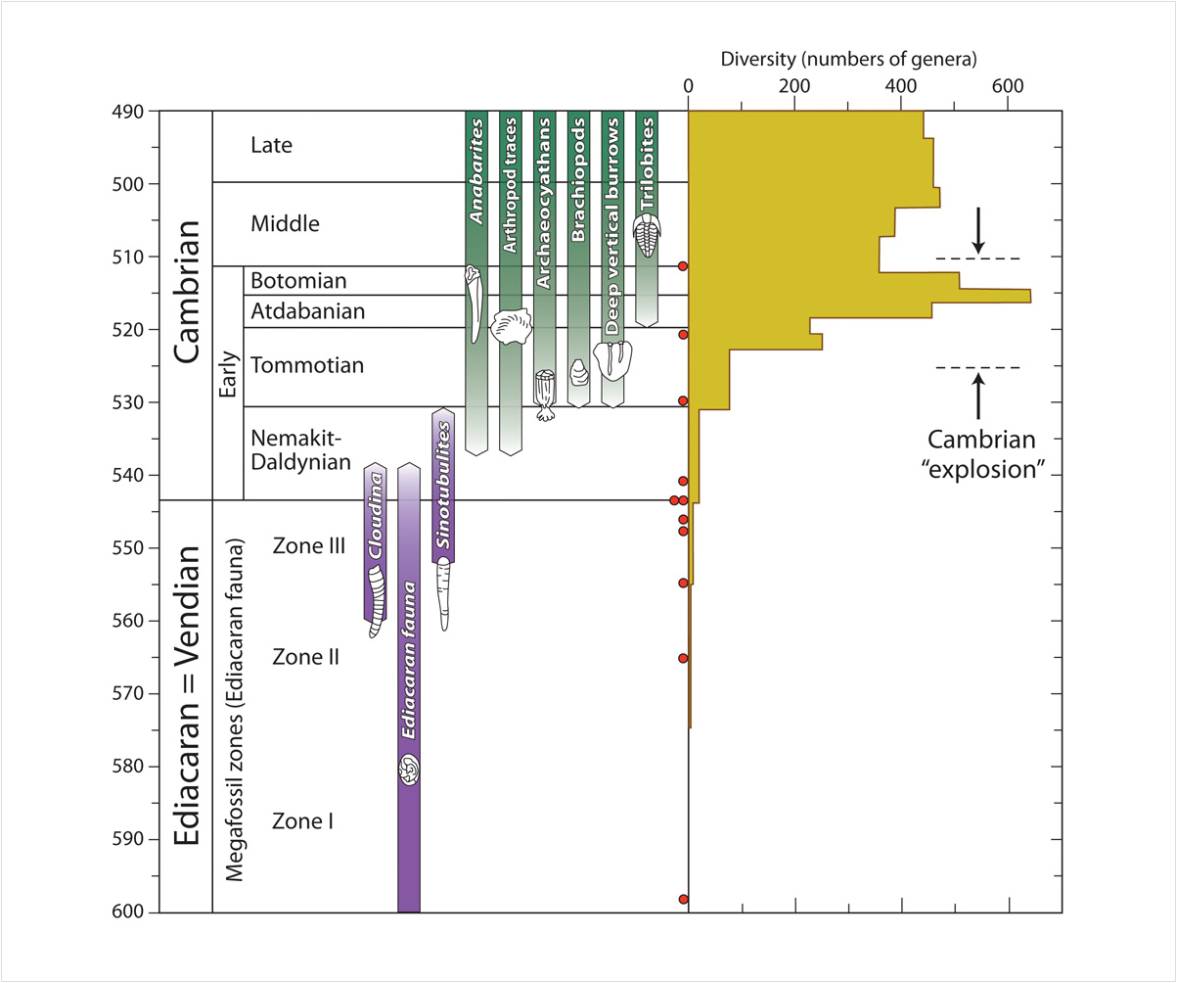
The big uptick in genera in the third (“Atdabanian”) stage of the Cambrian mainly comes from the fortuitous discovery of the huge Chengjiang lagerstätte. This figure debunks Meyer’s contention that essentially all significant fossilized animal phyla appeared in a 5-10 million year window. Prothero here is saying exactly what I wrote above, that Meyer misrepresents the fossil record by blanking out nearly all mention of the first 20 million years of the Cambrian. Prothero is also correct in writing:
Meyer takes the normal scientific debates about the early conflicts about the molecular vs. morphological trees of life as evidence scientists know nothing, completely ignoring the recent consensus between these data sets. Like all creationists, he completely misinterprets the Eldredge and Gould punctuated equilibrium model and claims that they are arguing that evolution doesn’t occur–when both Gould and Eldredge have clearly explained many times (which he never cites) why their ideas are compatible with Neo-Darwinism and not any kind of support for any form of creationism.
However, in his passionate outpouring, Prothero did not think through every statement carefully. This gave Casey Luskin, in his response, a few legitimate grounds for faulting Prothero. Luskin’s main attack on Prothero, however, is just rhetorical sleight of hand. Prothero wrote, “We now know that the ‘explosion’ now takes place over an 80 m.y. time framework.” He is, of course, not denying that many phyla first appeared in the fossil record in a 5-10 million year period. He is stating (accurately) that there was a lot else known to be going on before and after that narrow period, and of course that the first appearance of a species in the fossil record does not mean it did not exist earlier. But Luskin, by pulling up quotes from other geologists that define the Cambrian explosion more narrowly, simultaneously portrays Prothero as a misinformed Cambrian-explosion-denier and deflects attention from the crucial first 20 million years of the Cambrian.
A list of critical reviews of Darwin’s Doubt is given here by John Pieret. Many of these are on blogs or specialty sites like Panda’s Thumb. Two critical reviews that appeared in the more or less popular press are those by John Farrell in National Review, and by Gareth Cook in The New Yorker. Charles Marshall wrote a brief critical review in Science, which we discuss below.
“Darwin’s Doubt Critical Reviews” is a whole blog dedicated to the subject. There you can find the text of Farrell’s and Marshall’s reviews, as well as additional scholarly essays on the subject. John Harshman gives chapter by chapter technical objections, written in a tone of gentle irony. Harshman calls out Meyer’s misrepresentations around phylogenetics and the animal tree of life, and has this to say about the Douglas Axe research on which Meyer relies to prove the impossibility of evolution producing new genes:
[Axe] realizes that building a new organism requires building new proteins, apparently forgetting his claimed expertise in gene regulation, or perhaps only forgetting what promoters are. Oddly enough, this is followed fairly quickly by a citation of one of Ohno’s papers, of which the major point was that most Cambrian explosion animals had the same genetic tool kit, “nearly identical genomes, with differential usage of the same set of genes accounting for the extreme diversities of body forms.” But of course some of those nearly identical genes were new before the explosion, and that’s what Meyer wants to notice…
Next we discuss protein folds. How different does a tertiary structure have to become before it can be called a new fold? I have no idea, and Meyer doesn’t say. Here’s another thing Axe knew, because he was a protein scientist and they know stuff: “…new protein folds could be viewed as the smallest unit of structural innovation in the history of life.” And “…the ability to produce new protein folds represents a sine qua non of macroevolutionary innovation.” I guess Ohno was just kidding. So, having reduced macroevolution to the evolution of new protein folds, Axe finds that randomly replacing 1/5 of the exterior amino acids in a protein make it no longer functional, at least in its old role. We don’t know if it had a new function, but of course it’s hard to test for some unknown function. Axe also finds, surprisingly, that changes to a protein resting on an adaptive peak tend not to be selectively advantageous.
But here’s the important bit: Axe’s experiments show that it’s impossible (that is, so improbable as to have a low chance of ever happening, anywhere, during the entire history of life) for one functional protein fold to evolve into another, either gradually through selection or drift, or by macromutation. Thus the duration of the Cambrian explosion is irrelevant. The smallest unit of structural innovation is unable to emerge no matter how much time you give it. No new proteins can evolve. And macroevolution is all about new proteins. Oh, and “new function” is synonymous with “new fold”, so no new functions, ever. [But this is contradicted by post-Cambrian observations or inferences of new functions and genes being evolved, so Axe’s conclusions cannot be correct].
In another of the articles at the Darwin’s Doubt Critical Reviews blogspot, Elizabeth Liddle explains, with many diagrams, why Meyer’s characterization of the patterns in the fossil record is fundamentally mistaken. Meyer argued (Chapter 2):
According to Darwin’s theory… one would expect small-scale differences or diversity among species to precede large-scale morphological disparity among phyla…[However] the actual pattern in the fossil record, however, contradicts this expectation … Instead of more species eventually leading to more genera, leading to more families, orders, classes and phyla, the fossil record shows representatives of separate phyla appearing first followed by lower-level diversification on those basic themes.
Liddle replies,
Well, of course it does, Dr Meyer! You have just, in Chapter 2 of your fat book made an absolutely fundamental error of understanding of the entire principle of phylogenetics and taxonomy. No, of course you wouldn’t expect phyla to follow “lower-level diversification on those basic themes”. How could it possibly? And how could you possibly so fundamentally misunderstand the entire point of Darwin’s tree and its relationship to the nested hierarchies observe by Linnaeus?
All branching events, in Darwin’s proposal, whether the resulting lineages end up as different phyla or merely different species, start in the same way, with two populations where there once was one, and a short morphological distance between them. It is perfectly true that the longer both lineages persist for, the greater the morphological distance will become. But that isn’t because they started different, or because the phyla come later. It’s because what we call phyla are groups of organisms with an early common ancestor, whose later descendents have evolved to form a group that has a large morphological distance from contemporary populations who descended from a different early common ancestor.
So when a phylum, or a class, or even a kingdom first diverges from a single population into two lineages, the “morphological distance” from the other lineage will be very short. We only call it a “phylum” because eventually, owing to separate evolution, that distance [later] becomes very large.…So of course, if we look at the fossil record as these speciation-events were happening and try to categorize the organisms in terms of their modern descendents, we will find a great number of different phyla, and far fewer species.
The “Smilodon’s Retreat” author is laboriously writing chapter-by-chapter critiques of Darwin’s Doubt. At this point, he is only through Chapter 2 out of 20. It will be interesting to see how he engages with the genetic information arguments in the later chapters of the book. Larry Moran has finally bestirred himself and entered the fray in September-October with a barrage of posts on his Sandwalk blog that address most of the specific attacks by Meyer on mainstream molecular biology. This post summarizes many of these issues, and has links to the related posts. Moran deals with Meyer’s objections about assuming constant mutation rates, about different genes evolving at different rates, about discrepancies between different dating methods, and about the alleged circular reasoning involved in assuming common ancestry.
It is worth skimming down through the Comments on these Sandwalk posts. Often Moran’s main posts are fairly short, but knowledgeable commenters like Rasmussen, Harshman, and Diogenes respond to hostile questions and add relevant information and links to literature. For instance, in response to Meyer’s statement that “Any plausible ancestor to such organisms would have likely left some hard body parts, yet none have been found in the Precambrian,” I found these comments to be useful:
[It is] a strange thing to see that Meyer wants there to be hard body parts in the fossil record before they evolved. How…does that even make sense?
and
[Meyer] uses the old argument that some body plans make no sense without hard parts. This encapsulates a misunderstanding of the difference between “hard” and “mineralized”. Chitin is hard but not very preservable. Calcite is both. Most of the Burgess fossils have chitin (or some sort of cuticle) but no mineralization, and thus would be preserved only in exceptional circumstances.
The Discovery Institute, of course, relishes this controversy, and has published numerous articles seeking to refute these critical reviews. A list of these responses as of September 9 can be found here at the Discovery Institute’s Evolution News and Views site.
Larry Moran has offered a pithy evaluation of this set of responses; scroll down in the Comments on this post to find rebuttal of the DI’s fantasy that all scholarly or popular articles touching on the Cambrian published since June, 2013 are motivated by the need to respond to Darwin’s Doubt.
Matzke’s Early Critical Review
On June 19, the day after the book issued, then-graduate-student (now Dr.) Nick Matzke published a lengthy rebuttal on the Panda’s Thumb site. This is probably the single most influential review of Darwin’s Doubt. The National Review and New Yorker reviews drew on Matzke’s article, and the DI writers have found it necessary to devote at least four responses to Matzke. For the rest of my article here, I will focus on mainly this post, and on the DI response.
Matzke had worked for three years at NCSE before heading off to graduate school, so he was already familiar with Intelligent Design arguments. After six years in graduate school, he is a published technical expert in phylogenetics, far more knowledgeable than anyone at the DI. I cannot in limited space do full justice to the content. I will just point out some aspects that happen to stand out to me.
In order to understand some of what Matzke writes, you need to grasp the concept of crown-groups and stem-groups. This Wikipedia article explains it in about two pages. To oversimplify, imagine a deciduous tree like an oak or maple, with a trunk, and limbs off the trunk and further branches off the limbs. A crown-group might roughly correspond to say the top half of the tree, with a stem-group corresponding to a limb that branched off the trunk (stem) below the crown.
In Darwin’s Doubt, Meyer focuses almost entirely on the end of the Early Cambrian, i.e. the Atdabanian stage (c. 521-514 Mya), the time of the most rapid appearance of animal phyla in the fossil record. By presenting the appearance of “phyla” in a simple rake-type diagram, Meyer obscures key information. Within the groupings that we call phyla, considerably developmental change is evident in the fossils over the ages. It’s not like the first fossils of the phylum Chordata were full-fledged mammals or even modern-type fish. Also, the representatives of many animal phyla looked a lot like those in the other phyla in the early Cambrian. As more millions of years passed, the members of each phylum tended to become more differentiated from other phyla. This is in accord with expectations from evolution.
Biologos notes:
The major animal body plans that appeared in the Cambrian Explosion did not include the appearance of modern animal groups such as: starfish, crabs, insects, fish, lizards, birds and mammals. These animal groups all appeared at various times much later in the fossil record. The forms that appeared in the Cambrian Explosion were more primitive than these later groups, and many of them were soft-bodied organisms. However, they did include the basic features that define the major branches of the tree of life to which later life forms belong. For example, vertebrates are part of the Chordata group. The chordates are characterized by a nerve cord, gill pouches and a support rod called the notochord. In the Cambrian fauna, we first see fossils of soft-bodied creatures with these characteristics. However, the living groups of vertebrates appeared much later. It is also important to realize that many of the Cambrian organisms, although likely near the base of major branches of the tree of life, did not possess all of the defining characteristics of modern animal body plans. These defining characteristics appeared progressively over a much longer period of time.
As Matzke puts it:
… the earliest Cambrian relatives of the living phyla tend to be a lot more wormlike or sluglike than most modern representatives of the living phyla. Of course, many of the living phyla are basically still worms, and the more complex living phyla (e.g. molluscs, chordates) have early-diverging representatives or relatives that are rather more wormlike than the better-known representatives with more complex bodyplans. Even the earliest “fish” – actually either stem-group craniates, stem-group cephalochordates, or stem-group chordates – are basically filter-feeding worms that happen to swim. They don’t have jaws, scales, limbs, a bone skeleton, or anything else that most readers would associate with the word “fish”.
Matzke points out that Darwin’s Doubt withholds from the reader diagrams such as the two below, which synthesize much study of earlier Cambrian fossils, and show reasonable inferences of ancestral relations which undercut Meyer’s contention that all these animal forms appeared out of nowhere.

(source: Brysse 2008, originally Briggs & Fortey [Science, 1989], reproduced by Matzke)

(Source: Peterson et al., Paleobiology, 2005, reproduced by Matzke)
Skeptics of evolution may point out that scientists are simply assuming the existence of the common ancestors marked here as “range extension”. I would answer that objection with a statement and with a question. First, a statement: the assumption of common ancestry has, in general, worked well to explain and predict observations of fossils and genomic patterns. For instance, Neil Shubin predicted on the basis of evolutionary theory that a creature with features intermediate between fish and amphibians existed in rock layers of a certain age and depositional environment, and went excavating in the Arctic to find it. After five years of digging, he found the type of fossil (“Tiktaalik “) he expected. Where we do have reasonable amounts of fossils to work with (i.e. from the Cambrian onward), trees of common descent based on fossil morphology generally match those based on comparative genetics, so it is reasonable to extend this back to the Precambrian.
Second, a question: “What specific, concrete explanation do YOU propose for the appearance of the fossils whenever they do appear?” If the trilobites did not arise via evolution from some prior common ancestor, how did they come to exist? Man up and put your counter-proposal on the table. Did an Intelligent Agent create each genus or species of trilobite ex nihilo? Did the Intelligent Agent take pre-existing arthropods and miraculously rearrange their DNA? Well? Merely shouting “Design!” is worthless here. If you really believe that some particular miraculous intervention is how it happened, be honest and courageous enough to state that clearly.
Matzke points out that Meyer fails to present the broad sweep of observed changes in fossilized organisms, which are consistent with common ancestry and evolution. According to Matzke:
..we still have this sequence observed in the fossil record:
• 1. Before 700 mya, maybe well before: Single-celled eukaryotes (acritarchs)
• 2. Earlier Ediacaran: Multicellular animal eukaryotes, but simple, sponge-grade organisms
• 3. Later Ediacaran: Multicellular animal eukaryotes with more complexity, i.e. cnidarian-grade organisms
• 4. Very late Ediacaran: Simple slug-grade/worm-grade organisms (at least their tracks and burrows) – the first ones only making surface tracks and lacking burrowing ability. Making tracks suggests that the organisms have at least a front end and a back end, a mouth, anus, and gut connecting them. These are almost certainly bilaterians.
• 5. Very late Ediacaran: The very first biomineralized “skeletons”, e.g. Cloudina, basically a worm secreting a tube, as well as the first evidence of predatory boring. Cloudina gets no mention at all in Meyer’s book.
• 6. At the beginning of the Cambrian, we start to see more complex burrowing – e.g., vertical burrowing through sediment, clearly indicating worm-grade organization and an internal fluid skeleton, i.e. a coelom. The burrows gradually increase in complexity over 10 my.
• 7. Small shelly fauna: The shells, which started very small and very simple, gradually diversify and get more complex, radiating especially in the Tommotian. By the end of the Tommotian, some of the “small shellies” can be identified as parts of larger, “classic” Cambrian animals. The Tommotian is an utterly key period for any serious discussion of the Cambrian Explosion. Unfortunately, the word “Tommotian”, or any equivalent terminology (the detailed stratigraphy of the Cambrian is still being worked out, see Erwin & Valentine 2013 for a review), does not even appear in the book! The Small Shelly Fauna (SSF) gets just one (one!) mention in the book, buried in endnote 27 of Chapter 4, a whole chapter devoted to debunking the idea that the Ediacaran fauna is “ancestral” to bilaterians. …
• 8. The earliest identifiable representatives of Cambrian “phyla” don’t occur until millions of years after the small shelly fauna have been diversifying, and they tend to be taxa on the stem below the crown of living phyla, rather than placeable within the crown. Trilobites are an exception, but what is often missed is that deposits like the Chenjiang have dozens and dozens of trilobite-like and arthropod-like organisms that fall cladistically outside of these respective clades. These are transitional forms! How can this fact not be highlighted!?!
Matzke shows the following diagram, which illustrates many of the points he made verbally. There was a lot going on in the first 20 million years of the Cambrian, which is crucial to understanding the appearance of all the animal forms in the later Cambrian.

(Source: Marshall, Annual Review of Earth and Planetary Sciences, 2006, reproduced by Matzke)
(Note: this diagram uses the older nomenclature for the stages of the Cambrian period, such as “Tommotian” and “Atdabanian”; a diagram with similar information but using the current stage names appears here.) Matzke goes into some detail explicating various blunders in Darwin’s Doubt in the realm of phylogenetics. E.g., “[Meyer’s] main argument is basically that phylogenetic results sometimes conflict, therefore the whole thing is meaningless. This exhibits a jaw-dropping level of incompetence…Meyer just cites cases of phylogenetic conflict with reckless abandon…without any evidence of having any idea that causes of conflict are often well-known and can be predicted ahead of time,” and “But to creationists/IDists, all phylogenetic conflicts of any sort are considered equally, crashingly devastating. It’s rather a lot like when the young-earth creationists argued if estimates of the age of the Earth varied between 4.5 and 4.6 billion years ago, this 100-my disagreement was huge, and therefore we should instead think the Earth is 6,000 years old.” Matzke points out that there are objective mathematical tests that demonstrate that the current consensus trees of life are worthy of high confidence. Meyer’s treatment of Anomalocaris and “Lobopodia” come in for particular criticism.
Matzke writes:
The “key features” of the “crown groups” – groups defined as the clade containing two or more currently-living species, their common ancestor, and all of its descendants – did not come about all at once. Instead, when fossils are analyzed cladistically, we typically discover a bunch of species that morphological characters place below the crown – i.e., “stem groups”. The key thing about stem groups is that they have some, but not all, of the characters that make up the “body plan” of the crown clade under discussion. Among other things, this means:
…3c. The “body plans” did not originate instantaneously, all at once, unlike the ignorant contentions of creationists like Meyer.
3d. The “body plan” concept is basically just as arbitrary and flexible as the “phylum” concept – indeed, they basically both amount to a list of characters found in some monophyletic group (or, more commonly, a majority of the members of some phylogenetic group…
3e. The stem group taxa are the transitional forms that Meyer et al. are, allegedly, looking for.
The final section (“Other Issues”) of the body of Matzke’s review addresses the origin of genetic information:
Meyer’s main argument is really about “information”. Back in his Signature in the Cell book, Meyer asserted that the only known source of information was intelligence, and that therefore we could safely infer that intelligent design was behind the origin of life. This is problematic for all sorts of reasons, but one of the biggest was that intelligence is not the only known source of information – in particular, evolutionary processes of mutation+selection can produce it, thus intelligence is not the only cause of information, thus “information” isn’t some magical signal uniquely evidencing intelligence in the complete absence of any other evidence.
This section of Matzke’s review runs to some twenty paragraphs, and succinctly answers a number of the specific claims in Darwin’s Doubt that evolutionary processes cannot produce the genetic information required to account for the proliferation of animal life in the Cambrian. In answer to Meyer’s objection that academic papers assume the existence of an ancestral gene, Matzke responds:
Never mind that, when it comes to the Cambrian bilaterians, we have oodles of evidence for prexisting animals (cnidarians and sponges, both of which have lots of genes shared with other animals) as well as single-celled eukaryotes, which have plenty of genes, further back…Never mind that in any process of tracing a gene’s history back in time the trail will eventually go cold – it is a necessary result of mutational decay of sequence similarity – and this is not evidence of any miraculous process at the beginning of it, just evidence that human knowledge is limited.
…there’s no evidence that new protein domains were required in the Cambrian – I’d be surprised if any protein domains are known that are both unique to and required for the existence of Animalia. Animal genomes mostly just elaborate on and expand (through duplication) the already-complex pre-existing eukaryote genome… The basic protein domains mostly originated very far back in evolutionary history, where things like ultrahigh population sizes help out… [Note Matzke is talking about protein domains here, not whole proteins or genes].
And this:
Elsewhere in the book Meyer invokes some other arguments to justify the “improbability of functional sequence” argument. These are Behe & Snoke’s 2004 argument about multiple simultaneous mutations, Behe’s Edge of Evolution argument about “chloroquine complexity clusters” and protein-protein binding sites, and work by Douglas Axe and Anne Gauger, run out of the Discovery Institute’s approximately 2 person research institute, the Biologic Institute, which typically involves making evolutionarily absurd modifications to proteins and then showing that they don’t work. …Anyway, most of this has been rebutted elsewhere on PT [Panda’s Thumb], and there is little point in doing it again.
And this:
The multiple-required-mutations stuff, by the way, is basically just Behe’s refuted “irreducible complexity” argument disguised as an argument about sequence evolution, and is only relevant if it can be shown that 2 or more neutral mutations ever were required for anything relevant to the Cambrian Explosion, but, as is typical in DI literature, this is just blithely assumed rather than argued for. Showing it for any case would be nontrivial, and every detailed study I have read about multiple-mutation adaptations indicates that it doesn’t usually apply – instead, what typically occurs to produce an alleged “multiple mutations required” adaptation is that a variety of single-step mutations are selected as partial, imperfect adaptations to some chemical or environmental stressor. One or more of these enables the viability of some mutation of high adaptive value, which is then followed by selection of further mutations ameliorating whatever negative impacts the “major” adaptive mutation might have incurred. At the end of this process, you have something that would look as if several mutations would have had to happen at once, but only if you are ignorant of the somewhat circuitous-but-adaptive path (sometimes with selection for functions other than the final one) that was actually taken.
Luskin Contra Matzke
Casey Luskin is a writer for the Discovery Institute. Luskin has taken some graduate courses in biology, and is professionally trained as an attorney. Luskin quickly struck back with a piece entitled ”Rush to Judgment: Nick Matzke’s Hasty Review of Darwin’s Doubt Makes Bogus Charges of Errors and Ignorance.”
Luskin first tries to make a case that Matzke did not actually read the book before writing his review. This charge is ridiculous – Matzke could not have written in the detail he did, citing footnotes, etc., without having read the book, and Matzke has elsewhere described his timeline of reading the book and writing his review – – but that hasn’t stopped ID advocates from continuing to repeat this calumny in an effort to discredit Matzke.
Luskin completely ignores Matzke’s devastating expose’ of Meyer’s misrepresentation of the fossil record, especially Meyer’s blackout of the fossil evidence in the 20 million years immediately preceding the core Cambrian explosion. Not a word from Luskin on this.
Luskin issues a general defense of Meyer’s treatment of phylogenetics: “Meyer explains and critiques attempts to reconstruct phylogenetic trees based upon the comparisons of anatomical and genetic characters in his fifth and sixth chapters… Meyer critiques many of the assumptions and methods of cladistics in the context of the larger evaluation of phylogenetic reconstruction that he (Meyer) offers in those chapters…” Yes, Meyer did all these things, but one should re-read Matzke’s review to decide whether Meyer did these things well or poorly. Luskin devotes considerable space to defending Meyer’s treatment of Anomalocaris and “Lobopodia”, which I discuss below.
Luskin states that the relative suddenness of the appearance of many phyla in the Cambrian is only one aspect of Meyer’s critique of neo-Darwinism. The other, more central challenge is “that the neo-Darwinian mechanism lacks the creative power to produce the new animal forms that first appear in the Cambrian period.” According to Luskin,
Matzke scarcely addresses Meyer’s second and more central critique of neo-Darwinism. Matzke does not respond in any detail to any of Meyer’s multiple challenges to the creative power of the mutation/selection mechanism… Meyer offers five detailed scientific critiques of the alleged creative power of the mutation/selection mechanism. Yet Matzke in his nearly 10,000-word review offers no detailed response to any.
This is grossly misleading. A reader of Luskin’s piece would come away with the conviction that Matzke had no engaged this issue in any meaningful way. It is a common ID tactic to only offer partial truths. It is true that Matzke did not address every point in Darwin’s Doubt; to do so might require an equal-length tome. However, as noted above, Matzke explicitly acknowledged that Meyer’s chief argument regarded “information”, and Matzke did write some twenty paragraphs on the subject, and did address specific arguments in Meyer’s book. As I already cited above, Matzke stated explicitly why he did not devote more space to Meyer’s genetics arguments: ” …most of this has been rebutted elsewhere on PT [Panda’s Thumb], and there is little point in doing it again.”
In some cases Matzke dismissed a given argument with a single paragraph. In Chapters 10 (“The Origins of Genes and Proteins”) and 12 (“Complex Adaptations and Neo-Darwinian Math”), Meyer relies heavily on work by Douglas Axe to argue that evolution is unable to generate significant new genes. While others have exposed the fundamental flaws in Axe’s work, Matzke does it in one sentence: “Work by Douglas Axe and Anne Gauger… typically involves making evolutionarily absurd modifications to proteins and then showing that they don’t work.”
Matzke points out that, as best we can tell, not a lot of new protein domains needed to be evolved in the Cambrian. This undercuts Meyer’s core argument that enormous new genetic engineering was needed to produce the Cambrian fauna. We know that bilaterian animals were well-established before the Cambrian even began, and even those animals use genes similar to those found in earlier, microscopic eukaryotes. Even after some 600 million years of divergence, humans still share nearly 70% of their genes with the lowly sponge.
Matzke’s Reply to Luskin
Whilst sitting in an airport waiting for his plane, Matzke banged out a response (“Luskin’s Hopeless Monster”). He explained that he had read Darwin’s Doubt the day it came out, and wrote his big review the next day. He has been living and breathing phylogenetics for years, as well as being a veteran of countering ID, so it was feasible for him to smoke the keyboard and write his long review in less than 24 hours.
Matzke corrects a key misrepresentation in Luskin’s piece. Luskin had written:
Matzke does attempt to address the first problem posed by the Cambrian explosion. He does so by claiming that methods of phylogenetic reconstruction can establish the existence of Precambrian ancestral and intermediate forms – an unfolding of animal complexity that the fossil record does not document.
Here Luskin is playing the same game that Meyer did in Darwin’s Doubt, of deflecting attention from the early Cambrian to the Precambrian. Matzke responds:
Well, no. I claimed that phylogenetic methods can establish, and have established, the existence of Cambrian intermediate forms, which are collateral ancestors of various prominent living phyla. The case is clearest with the most common and most-fossilized Cambrian phylum, the arthropods, but there is a fair bit of similar evidence for other major phyla. (Some phyla, primarily soft-bodied worms, have few fossils anyway, and there of course intermediate fossils are scare, although even if we had them they would be difficult-to-identify worms.) All of the leading authorities (Valentine, Erwin, Conway Morris, Briggs, Budd, etc.) would agree with me. More precisely, I agree with them, and they have all said in print what I just said. Furthermore, they would all agree that this is extremely important evidence for understanding the origin of “phyla”, evidence which cannot be ignored. But Meyer/Luskin ignore it, instead occupying themselves with hunting around in the Precambrian.
Similarly, Valentine, Erwin, Conway Morris, Briggs, Budd, etc., would all agree that it is utterly impossible to have a sensible discussion of the Cambrian Explosion while ignoring the 30-million year sequence of surface-crawling worms, then burrowing worms, then armored worms, then small shellies, THEN identifiable relatives of phyla, most of which are (not coincidentally) stem groups rather than members of the crown phyla, and which have characters suites transitional between the major crown phyla. These are fatal, catastrophic omissions Meyer’s book, which is allegedly supposed to be a serious commentary on the Cambrian Explosion.
Fossils of Anomalocaris (“anomalous shrimp”) and the Lobopodia group share some but not all characteristics with some well-established phyla. As such, they are exactly the sorts of intermediates that support the evolutionary interpretation of the fossil record, a point avoided or denied by Meyer. It is important to distinguish between crown and stem groups, which Meyer did not do sufficiently. Matzke noted this in his original critique, and then at considerable length in his response here to Luskin. But Luskin, in his piece, writes (inaccurately) as though the issue was whether or not Meyer made a factual error in referring to Anomalocaris as an arthropod and Lobopodia as a phylum. This allowed Luskin to apparently vindicate Meyer by citing various other authorities who (like Meyer) refer to Anomalocaris an arthropod and Lobopodia as a phylum, while side-stepping Matzke’s substantive objections on the subject.
In late October, Matzke published a follow-up article on Panda’s Thumb, addressing claims by Discovery Institute writers that the critics aren’t addressing Meyer’s arguments. This article is a mainly a tighter, updated version of Matzke’s original article. It includes a nice summary, with graphics, of Legg, et al. (2013) on arthropod ancestral relations.
Charles Marshall’s Review in Science
In the September 20, 2013 issue of Science, Harvard paleontologist Charles Marshall published “When Prior Belief Trumps Scholarship,” a brief review of Darwin’s Doubt. Marshall’s article was critical but fair:
Meyer’s scientific approach is negative. He argues that paleontologists are unable to explain the Cambrian explosion, thus opening the door to the possibility of a designer’s intervention. This, despite his protest to the contrary, is a (sophisticated) “god of the gaps” approach. Despite its readability and a plethora of scholarly references, however, there are substantial omissions and misrepresentations. For example, Meyer completely omits mention of the Early Cambrian small shelly fossils and misunderstands the nuances of molecular phylogenetics, both of which cause him to exaggerate the apparent suddenness of the Cambrian explosion.
…His case against current scientific explanations of the relatively rapid appearance of the animal phyla rests on the claim that the origin of new animal body plans requires vast amounts of novel genetic information coupled with the unsubstantiated assertion that this new genetic information must include many new protein folds.
This is all true, and Marshall’s proposal for how the new phyla emerged is reasonable, although it is not the whole story:
In fact, our present understanding of morphogenesis indicates that new phyla were not made by new genes but largely emerged through the rewiring of the gene regulatory networks (GRNs) of already existing genes. Now Meyer does touch on this: He notes that manipulation of such networks is typically lethal, thus dismissing their role in explaining the Cambrian explosion. But today’s GRNs have been overlain with half a billion years of evolutionary innovation (which accounts for their resistance to modification), whereas GRNs at the time of the emergence of the phyla were not so encumbered.
On the one hand, it may be a good thing to publicize this perspective on the weakness of ID. An unintended consequence, however, is that this review gives the ID movement grounds to claim that Darwin’s Doubt is being taken seriously by the scientific establishment. Intelligent Design proponents promote the notion that science is, with each passing year, finding that “Darwinism” is inadequate or untenable. They crave controversy, since that gives them grounds to argue that schools should “teach the controversy,” instead of monolithically promoting unguided evolution.
To date, the critical reviews of Darwin’s Doubt have been mainly published in private or specialized blogs or sites that were already noted for defending evolution. The articles in National Review and The New Yorker were by authors with a history of opposing ID, and so were in that sense unremarkable. However, this recent review by a leading paleontologist, published in one of the most prestigious scientific journals in the world, can and will be used by supporters of ID to claim that Meyer’s work is threatening to the scientific establishment.
Here is one gleeful example I picked at random from a conservative Christian blog:
Distinguished UC Berkeley Paleontologist Reviews Meyer’s Book “Darwin’s Doubt” (a book too important to ignore)
UC Berkeley paleontologist Charles Marshall’s review of Stephen Meyer’s book Darwin’s Doubt: The Explosive Origin of Animal Life and the Case for Intelligent Design represents a significant turn of events. Finally, about three months after the publication Meyer’s book, a reputable Darwinist actually addresses the main evidence and arguments in Meyer’s case for ID from the Cambrian explosion. This review unwittingly demonstrates that Meyer’s book raises questions that the evolutionary establishment can’t simply ignore. This is precisely, at minimum, what Meyer had hoped to accomplish. Congratulations to my colleague Stephen Meyer and to his worthy debate partner, the distinguished Charles Marshall.
This blog post echoes the fiction that the DI writers keep recycling, namely that no critical review besides Marshall’s has actually addressed “the main evidence and arguments in Meyer’s case for ID.” That is a plain falsehood. Beside Matzke’s review which DID address the capability of evolution to generate new genetic information, I have linked above to articles by Harshman and by Matheson and by Moran which likewise directly answer the claims of Axe and of Meyer.
Another consequence of Marshall’s review is that it provides an august opportunity for Meyer himself to step forward and publish a series of four responses to Marshall, with as much fanfare as possible. Hitherto, the work of defending Darwin’s Doubt has fallen to the lesser employees at the DI.
Meyer’s first response was not very promising. There Meyer sought to respond to Marshall’s claim that gene regulatory networks (dGRNs), which are necessary for the development of animals, could have been more labile or flexible in the past. Meyer then proceeded to reiterate how inflexible such networks are today (which was never in dispute) and then he stopped dead, never engaging the literature which supports Marshall’s proposal that things may have been different some 600 million years ago than they are now.
In Closing
It is unlikely that many adult minds will be changed by all these reviews and rebuttals. Young people who take college-level biology courses will in general catch on that evolution is well-supported by the facts. Those who come from conservative religious backgrounds and who do not engage biology or geology will likely retain the opinion that they could not possibly have come from monkeys and the earth is actually only 6000 years old. I know a lot of those people – -they are not foolish on the whole, but the creationist mindset is hard to break free of. The communications put out by the DI are well-crafted to appeal to them. This probably doesn’t do much real harm; in light of recent court decisions, it is unlikely that ID will find its way into the schools. My main personal objection is that the ID movement promotes looking for God’s activity in the crevices of what is unknown, rather than celebrating His providence in what is known.
************************************************
For a brief description of related articles, e.g. review of John Sanford’s Genetic Entropy, discussion of Grand Canyon geology, etc., see ABOUT.

Pingback: Letters to Creationists
Wow, really great article. Thank you for your efforts here. I had read most of the reviews that you mention here and you have done a fantastic job of capturing the essence of those reviews. Another bit that I really jumped out at me what your comment about Ceolacanths. It struck me as a really powerful example of the fickle nature of the fossil record that I had not thought about.
Joel, thanks for the kind words. Hey, I really enjoyed your article on going to the Joggins fossil cliffs– a different sort of family vacation! ( http://thenaturalhistorian.com/2013/06/04/trip-to-joggins-fossil-cliffs-nova-scotia-polystrate-trees/ )
-Scott
Yeah, cool review!
Pingback: Evolution and Faith: My Story, Part 2 | Letters to Creationists
Hey Scott,
Great review, thanks!
A small correction: Meyer is a fellow of the Discovery Institute, not the Design Institute. The mistake appears multiple times in the article.
Shadi, thanks for the catch. Fixed!
Pingback: Realistic Expectations for Transitional Fossils | Letters to Creationists
It’s all “gross” theory. I hate to generalize and not give point for point rebuttal but after stepping back and looking at the assumptions – it is clearly interpretation based on worldview…. I’m amazed other Christians who are TE don’t see it. If the inductions are open to error then clearly all of the “actual” evidence could be interpreted quite differently. Question everything.
What amazes me is that the none of the people here seems to have compared Meyer’s Figure 3.8 with either Prothero’s claims about it, nor with Prothero’s and Matzke’s diagrams. And that includes the author of the long article, whose name I cannot find anywhere on this webpage. It even includes PROTHERO and MATZKE!
A cursory look at Meyer’s 3.8 shows that it agrees with Prothero’s diagram in showing the Cambrian to have begun 543 mya. But look: Meyer’s 3.8 shows the “Main Pulse of the Cambrian explosion” starting at 530 mya, and ending at 525 mya, which Prothero’s diagram puts in the Tommotian, and the shelly fossils have their explosion in this same time period in Matzke’s diagram.
And yet, Prothero alleged in his review that Meyer claims the same Figure 3.8 misleads readers into thinking that the animal phyla all erupted in the Atdabanian! And the author of the long article on which I am commenting is oblivious to all that I said about Meyer’s figure 3.8, yet makes the irrelevant complaint that Meyer nowhere names the Tommotian in his book — but then, Meyer nowhere names the Atdabanian either! The only part of the early Cambrian that Meyer names is the Mayikaian, in his endnote about shelly fossils.
There is much more amiss in the article, but this will do for a first installment.
Pingback: A Survey of Biblical Natural Theology | Letters to Creationists
Pingback: Exposing the Roots of Young Earth Creationism | Letters to Creationists
A really amazing review of Meyer’s book! Thank you! I don’t know when I’ve ever seen such a thorough analysis on a blog page. I also appreciate your theological perspective.
Thanks for the kind words — I hope to provide resources that help folks embrace the truth.
Pingback: A Creationist Speaker Comes to Town | Letters to Creationists
Pingback: Whatever Happened to Intelligent Design Theorist William Dembski? | Letters to Creationists
Pingback: Listing of Articles on Science, Faith and Other | Letters to Creationists
Pingback: Evolution Before Our Eyes: Complex Mutations in Microbes Giving New Functions | Letters to Creationists
Pingback: Science and Faith at the American Scientific Affiliation 2018 Meeting | Letters to Creationists
Seems to me there are all sorts of attempts to construct phylogenetic trees based on genetic drift and based on cladistics. They don’t converge to a clear answer. Whatever happened, there was only one reality.
Seems to me it is easy to slur an ID approach because the evolutionary paradigm is terminally flexible and such widespread orthodoxy that you can run down the assumptions of an ID protagonist as easily as you can run down someone with a conflicting take on details who subscribes to the host theory.
Seems to me there will be a very strong background pressure to uphold the neo-Darwinian paradigm simply because to admit doubt or failure is now unthinkable because of the consequences for pride, finances, credibility etc.For some your world would fall apart.
Seems to me we have very very poor basic understanding of genome. Witness we cannot read ‘blind’ entire genome into phenotype (complete organism). We just have snippets of understanding and a lot of hubris/hype. How then can we make confident assertions on genetic drift pathways?
Seems to me a lot of the defences of evolutionary paradigms are disingenuous. They allude to a solid orthodox understanding which doesn’t actually exist.
Seems to me Meyer’s reasoning in ‘Signature in the Cell’ is pretty sound, and the criticism unfocused and lacking in rigour.
Seems to me that all godbots are dishonest.
All this goes a fair way back by now and I think the arguments for ID are going considerably beyond the Cambrian Explosion now and are heading more in the direction of biology, mathematics and information theory. The Cambrian may be of utmost importance to paleontologists to maintain their shaky hold on Darwin, but more ordinary yet curious people would require a more summarized short form presentation of these breath-taking so-called facts to make sens of them. It appears something’s well amiss here.
Nicholas,
First, if you are like most ID supporters, you are an evangelical Christian who would like to be able to claim that your faith is supported by physical facts. As a fellow evangelical, please allow me to suggest that you look for God more in what we do know than in what we don’t know. Despite their strenuous denials, the ID spokesmen are engaging in nothing more than the old god-of-the-gaps approach (trying to find something we cannot yet explain, and claiming that it requires supernatural intervention, which they call “design”). That is looking for God in what we don’t know, and it ends up bringing disgrace on the Christian faith. St Augustine wrote long ago (430 A.D.) that if non-Christian men who are knowledgeable about the physical world hear Christians claiming that their BIble teaches things about nature which these men know to be nonsense, it will make it harder for these men to take seriously the truth-claims of the gospel.
Alternatively, we can point to evidence that we do know, namely the historical evidence for the life and especially the resurrection of Jesus of Nazareth. I wrote that up here: https://letterstocreationists.wordpress.com/historicity-of-jesus/ . The resurrection is the “sign” that Jesus himself pointed to, to authenticate his claims.
Getting to the Cambrian Explosion, here is a “summarized short form presentation” as you requested:
. . . . . . . . . . . . .
Meyer and his colleagues systematically suppressed lots of known facts that show that the appearance of various phyla in the fossil record was not nearly so sudden as claimed. Moreover, the particular specimens of each new phyla that appeared then were very different from modern species – – e.g., yes, representatives of “Chordata” show up in the Cambrian, but they are basically swimming worms with a nerve cord down their backs, not fish with backbones and jaws.
The key thing to note here is that we simply don’t have a continuous record of the fossils in the Cambrian/Precambrian — it was so long ago that nearly all of whatever fossil beds did form back then have long since been eroded or subducted or squashed/cooked beyond recognition; and before hard body parts evolved, only in the rarest of circumstances would any kind of fossil impression be formed at all. Of the modern phyla that are soft worms, we usually have zero fossil record. We only have a few good fossil sites that give snapshots of the fauna at some one particular site, maybe once every ten million years or so, during an era when there was rapid evolution [it seems there was a spike up in atm oxygen content around then]. Given that highly discontinuous sampling of the fossil record, OF COURSE the fossil record we have will *look* discontinuous.
. . . . . . . . . . .
I am aware that ID proponents make a lot of claims re information theory, but these claims have been thoroughly discredited by actual scientists (tho the ID proponents won’t admit it). See e.g. here https://letterstocreationists.wordpress.com/2017/11/04/evolution-before-our-eyes-complex-mutations-in-microbes-giving-new-functions/
and Randy Isaac’s presentation about halfway through this post, https://letterstocreationists.wordpress.com/2018/08/20/science-and-faith-at-the-american-scientific-affiliation-2018-meeting/ .
However, the issues are complex enough that if you don’t want to accept these rebuttals I cannot persuade you.
But that is getting off-topic from this post on Cambrian fossils.
Peace and blessings…
Pingback: Fazale Rana Disputes Evolution [2020 NCCA Apologetics Confc, 3] | Letters to Creationists
Pingback: Scientists’ Responses to Stephen Meyer’s Return of the God Hypothesis | Letters to Creationists
Pingback: 低端护教学(6)|启底年轻地球论(二) – Eddy & Emma's Blog